|
||||
|
|
Часть I ОБЩАЯ ХАРАКТЕРИСТИКА ПСИХИЧЕСКОЙ ДЕЯТЕЛЬНОСТИ ЖИВОТНЫХ Глава 1 ПРОБЛЕМА ИНСТИНКТА И НАУЧЕНИЯ Ранние представления о психической деятельности животных Начало познания поведения животных При изучении любой формы психической деятельности прежде всего встает вопрос о врожденном и индивидуально приобретаемом, об элементах инстинкта и научения в поведении животного. В широком плане это вопрос о внешних и внутренних факторах поведения, о его генетических основах и воплощении видотипичного поведения в индивидуальной жизни особи, о мотивации поведения. Еще древние мыслители, пытаясь постичь основы и движущие силы поведения животных, задумывались над тем, существует ли у животных какое-то «духовное начало», и если да, то какова его природа. Так, Эпикур и его последователи, особенно Лукреций, отвечали на этот вопрос положительно, но при этом со всей определенностью отстаивали положение о материальности такой «души». При этом Лукреций уже высказывал мысль, что целесообразные действия животных являются результатом своего рода естественного отбора, ибо выживать могут лишь животные, обладающие полезными для них свойствами. Древние философы-идеалисты исходили из представления о некоем изначальном «мире идей», «мировом разуме», аналогичном понятию Бога в более поздних церковных учениях. Порождением этого универсального «разума» является душа человека и животных, которая, по Сократу, соединившись с телом, подвергается влиянию чувствительности и направляется в своих действиях влечениями и страстями. Платон считал, что все поведение животных определяется влечениями. Отсюда был уже только шаг до представления об инстинктах. Аристотель, «величайший мыслитель древности», как его называл К. Маркс, был и первым подлинным естествоиспытателем среди философов. Человеку Аристотель приписал бессмертную «разумную душу» — воплощение божественного духа. Душа, по Аристотелю, оживляет тленную материю, тело, но только тело способно к чувственным впечатлениям и влечениям. Поэтому в отличие от человека, наделенного разумом, способностью к познанию и свободной волей, у животных имеется лишь смертная «чувственная душа». Но всем животным с красной кровью и рождающим живых детенышей присущи те же пять чувств, что и человеку. Поведение животных направлено на самосохранение и продолжение рода, мотивируется же оно желаниями и влечениями, ощущениями удовольствия или боли. Наряду с этим поведение животных определяется, по Аристотелю, и разумом, представленным у разных животных в разной степени. Разумных животных Аристотель считал способными к пониманию цели. Свои суждения Аристотель обосновывал конкретными, причем подчас поразительно точными, наблюдениями (наряду, правда, и с анекдотичными данными). У муравьев, например, он подметил зависимость их активности от внешних факторов (освещения). В отношении ряда видов животных он указал на их способность к научению друг у друга; описал он также ряд случаев звукового общения животных, особенно в период размножения. Пожалуй, впервые именно у Аристотеля мы встречаем ссылки на экспериментальные данные о поведении животных. Так, он указывал на то, что после удаления птенцов от родителей они научаются петь иначе, чем последние, и отсюда выводил заключение, что способность к пению не является «даром природы». Таким образом, уже Аристотель обосновывает представление об индивидуальном приобретении определенных компонентов поведения. Эту способность к индивидуальному научению и к запоминанию выученного Аристотель признавал за многими животными и придавал ей большое значение. Ряд положений Аристотеля получил дальнейшее развитие в учениях стоиков, хотя в некоторых отношениях здесь обнаруживаются и существенные расхождения. У стоиков впервые появляется понятие инстинкта (horme — греч., instinctus — лат.). Инстинкт понимается этими философами как прирожденное, целеустремленное влечение, направляющее движения животного на приятное, полезное и уводящее его от вредного и опасного. Хризипп указывал в этой связи на то, что если утята были выведены даже курицей, их тем не менее влечет к родной стихии — воде, где им обеспечено питание. В качестве других примеров инстинктивного поведения Хризипп ссылался на гнездостроение и заботу о потомстве у птиц, постройку сот у пчел, умение паука ткать паутину. Все эти действия совершаются животными, по Хризиппу, бессознательно, без участия разума (которого у животных нет), без понимания своей деятельности, на основе чисто врожденных знаний. При этом Хризипп отмечал, что подобные действия выполняются всеми животными одного и того же вида одинаково. Таким образом, Хризипп предвосхитил в ряде существенных моментов современный научный взгляд на поведение животных. Аналогично и Сенека-младший считал, что животные обладают врожденным знанием того, что им вредно, например врожденным знанием внешнего вида хищников. При этом Сенека также указывал на однообразие форм и результатов прирожденной деятельности животных. Четко разграничивая врожденное и благоприобретаемое поведение, Сенека отрицал наличие у животных разума, способности к мышлению. Инстинкт же в концепциях стоиков — властный зов Природы, которому животное вынуждено следовать безоговорочно, «не рассуждая». Как мы видим, уже в трудах античных мыслителей выступали во всей своей сложности основные проблемы поведения животных: обсуждались вопросы врожденного и благоприобретаемого поведения, инстинкта и научения, а также роль внешних и внутренних факторов психической активности животных. При этом выявлялись те в корне различные подходы к изучению психики животных, которые определяли дальнейший научный поиск в этой области, а именно материалистическое и идеалистическое понимание сущности психической деятельности. Доэволюционные взгляды на поведение животных (XVIII в.) После тысячелетнего застоя научной мысли во времена средневековья началось возрождение научного творчества, но только в XVIII в. делаются первые попытки изучения поведения животных на прочном фундаменте достоверных фактов, получаемых в результате тщательных наблюдений и экспериментов. В середине и конце этого века появляются труды целой плеяды выдающихся ученых, философов и натуралистов, оказавших большое влияние на дальнейшее изучение психической деятельности животных. Из них необходимо прежде всего упомянуть французского философа-материалиста, врача по образованию Ж.О. Ламеттри. Ламеттри придерживался взгляда, что инстинкты животных представляют собой совокупность движений, выполняемых вынужденно, вне зависимости от размышлений и опыта. При этом он подчеркивал биологическую приспособленность инстинктов, хотя и не мог определить причину этого. Исключительно важной чертой исследований Ламеттри являлся сравнительный подход к изучению психической деятельности животных. Сопоставляя психические способности разных млекопитающих, а также птиц, рыб, насекомых, он показал прогрессивное усложнение этих способностей по направлению к человеку. Осталось сделать только шаг к представлению об историческом развитии психики. Взгляды Ламеттри, сформулированные им на основе тогдашних знаний об анатомии и физиологии нервной системы, оказали впоследствии большое влияние на научное творчество Ламарка. Крупный французский просветитель Э. Б. Кондильяк в своем «Трактате о животных» (1755) специально рассматривал вопрос о происхождении инстинктов животных. Исходя из сходства инстинктивных действий с действиями, выполняемыми по привычке, Кондильяк пришел к выводу, что инстинкты произошли из разумных действий путем постепенного выключения сознания: разумное поведение превращалось в привычку, а последняя — в инстинкт. Против такой трактовки категорически возражал Ш. Ж. Леруа. Этот натуралист и мыслитель доказывал, что обозначенный Кондильяком ряд следует читать как бы в обратном порядке. В опубликованных им в 1781 г. «Философских письмах об уме и способности животных к совершенствованию» он выдвигает задачу изучения происхождения разума от инстинкта животных в результате повторяющегося действия ощущения и упражнения памяти. Эту идущую вразрез с церковными догмами концепцию развития высших психических способностей Леруа пытался обосновать и собственными данными о поведении животных в природе. Леруа придавал особое значение полевым исследованиям и настойчиво доказывал, что психическая деятельность животных и особенно их инстинкты могут быть познаны лишь при наличии всесторонних знаний об их естественном поведении и с учетом их образа жизни. Леруа видел в инстинктах животных воплощение их потребностей: необходимость удовлетворения последних и приводит к появлению инстинктов. Привычки, по Леруа, могут передаваться по наследству и в результате включаться в естественный поведенческий комплекс. Леруа иллюстрировал это на примерах охотничьих собак, передающих свои повадки потомству, или кроликов, перестающих рыть норки после того, как несколько поколений их жило в домашних условиях. Выше уже упоминалась исключительная роль Ж. Л. Л. Бюффона в формировании зоопсихологии как науки. Бюффон не только подошел к изучению психической деятельности животных как истинный натуралист, но и сумел при анализе своих полевых наблюдений и опытов воздержаться от антропоморфных толкований поведения животных (в отличие от многих ученых конца XIX и начала XX в.). Согласно его учению, животным (конкретно имелись в виду млекопитающие) свойственны разные формы психической деятельности, в частности ощущения и привычки, но не понимание смысла своих действий. Равным образом животные способны общаться, но их язык выражает лишь чувственные переживания. Выявляя зависимость между особенностями психической деятельности животных и их образом жизни, Бюффон выдвинул тезис о связи воздействий окружающей среды с внутренним состоянием («желаниями») животного, видя в этом определяющий фактор его поведения. Примечательными являются и его выводы о приспособительном значении психики, а именно о том, что психические качества животных являются не менее значимыми для сохранения видов, чем физические качества. Все концепции Бюффона, построенные на реальных фактах, вошли в созданную им единую систему естествознания и стали первой основой будущей науки о поведении и психике животных. Нам здесь важно отметить, что подлинно научное изучение инстинктов животных (начатое Бюффоном раньше Леруа) привело его к заключению, что сложные действия животных являются результатом сочетания врожденных природных функций, доставляющих животному удовольствие, и привычек. Тем самым Бюффон предвосхитил современное понимание структуры поведения животных, намного опередив своих современников. Проблема инстинкта и научения в свете эволюционного учения Ж. Б. Ламарк К началу XIX в. проблема инстинкта и связанный с ней вопрос о взаимоотношениях врожденных и приобретаемых действий животных привлекают к себе все большее внимание. Интерес к этим вопросам был обусловлен зарождением идеи трансформизма, появлением первых эволюционных теорий. Актуальной задачей стало выявление того, что передается в поведении по наследству в «готовом виде», что формируется в результате воздействия среды, что является всеобщим, видовым, а что — индивидуально приобретаемым, каково значение разных компонентов поведения в процессе эволюции, где проходит грань между человеком и животным и т. д. Согласно метафизическим взглядам на природу, господствовавшим на протяжении столетий, инстинкты животных представлялись незыблемыми, пребывающими в неизменном состоянии с момента их «сотворения». Творцам эволюционных теорий предстояло объяснить происхождение инстинктов и на конкретных примерах показать их изменчивость, как они это делали и в отношении видотипичного строения животного организма. Первое целостное учение об эволюции живой природы было сформулированно в самом начале XIX в. Жаном Батистом Ламарком. Как известно, Ламарк положил в основу своей эволюционной концепции представление о направляющем действии психического фактора. Он полагал, что внешняя среда действует на животный организм опосредованно, путем изменения-поведения животного. Вследствие этого опосредованного влияния возникают новые потребности, которые в свою очередь влекут за собой изменения в строении организма путем большего упражнения одних и неупражнения других органов, т. е. опять-таки через поведение. При всей ошибочности общих положений этой концепции (примат психики как некоего изначального организующего фактора, стремление организмов к «совершенствованию» и т. д.) великой заслугой Ламарка остается то, что он указал на огромную роль поведения, психической деятельности в процессе эволюции. Стремясь к объяснению всего исторического развития органического мира естественнонаучным путем, Ламарк доказывал, что даже самые сложные проявления психической деятельности развились из более простых и должны изучаться именно в сравнительно-эволюционном плане. При этом он со всей определенностью отрицал существование какого-то особого духовного начала, не связанного с физическим строением животного (как и человека) и не поддающегося естественнонаучному изучению. Все без исключения психические явления Ламарк признавал теснейшим образом связанными с материальными структурами и процессами и поэтому считал их вполне познаваемыми опытным путем, идя по общему пути естественнонаучного познания мира. Особое значение Ламарк придавал при этом связи психики с нервной системой. В противоположность концепциям психофизического параллелизма, которые бытуют и среди ряда современных этологов, Ламарк убежденно отстаивал положение о функциональной связи психики с нервной системой, о зависимости психической деятельности от нервной деятельности, о зависимости психической эволюции от эволюционных преобразований в нервной системе. Можно с полным основанием сказать, что Ламарк заложил основы сравнительной психологии, сопоставляя строение нервной системы с характером психической деятельности животных на разных уровнях филогенеза. С этих позиций Ламарк подходил и к проблеме инстинкта. «….Инстинкт животных, — писал он, — это наклонность, влекущая, вызываемая ощущениями на основе возникших в силу их потребностей и понуждающая к выполнению действий без всякого участия мысли, без всякого участия воли».[12] При этом Ламарк не считал инстинктивное поведение животных чем-то раз и навсегда изначально данным и неизменным. По его представлениям, инстинкты возникли в процессе эволюции в результате длительных воздействий на организм определенных агентов среды. Эти направленные действия и привели к совершенствованию всей организации животного через формирование полезных привычек, которые закрепились в результате многократного повторения, ибо такое повторное выполнение одних и тех же движений приводило к проторению соответствующих нервных путей и все более легкому прохождению по ним соответствующих нервных импульсов («флюидов», по терминологии Ламарка). Итак, по мнению Ламарка, тождественная инстинкту наклонность «стоит в связи с организацией, преобразованной привычками в благоприятном для нее направлении, и возбуждается впечатлениями и потребностями, движущими внутреннее чувство особи и предоставляющими ему случай послать — в связи с требованием действующей наклонности — нервный флюид к подлежащим мышцам».[13] Как отмечал академик В. Л. Комаров, Ламарк понимал под флюидами не что-то мистическое, а вполне материальное начало, близкое к современному пониманию энергии. «…Привычка упражнять тот или другой орган или ту или другую часть тела для удовлетворения вновь и вновь возрождающихся потребностей, — продолжал Ламарк, — делает для нервного флюида… движение в направлении органа, связанного со столь частым употреблением его, настолько легким, что данная привычка становится как бы неотъемлемым природным свойством особи».[14] Потребности порождают привычки (т. е. обусловливают процесс научения), привычки же превращаются в наклонности, которым животные «не могут противостоять, в которые они сами по себе не в состоянии внести никаких изменений. Отсюда ведут свое начало их привычные действия и их частные наклонности, которым дано название инстинкта». Вот как Ламарк представлял себе взаимоотношения между инстинктом и научением: «начало инстинкта в животных» — происхождение инстинктивных действий посредством пожизненного видоизменения двигательной активности. Необходимо еще добавить, что, по Ламарку,«наклонность животных к сохранению привычек и к повторению связанных с ними действий… распространяется затем в особях путем воспроизведения, или размножения, сохраняющим организацию и расположение частей в достигнутом состоянии, так что одна и та же наклонность уже существует в новых особях раньше, чем они начали упражнять ее».[16] Такая передача инстинктов от поколения к поколению «без заметных отклонений будет продолжаться до тех пор, пока не произойдет перемены в условиях, существенных для образа жизни».[17] В этих словах особенно отчетливо выражено представление об определяющей роли условий среды в эволюции инстинктивного поведения. Конечно, сегодня нам известно, что Ламарк ошибался, полагая, что переход индивидуально приобретенных в наследственно закрепленные формы поведения возможен путем многократного повторения выученных (или видоизмененных) движений, что привычки могут непосредственно обусловить генетические изменения организации животного организма. Итак, Ламарк видел в инстинктах животных не проявления какой-то таинственной сверхъестественной силы, таящейся в организме, а естественные реакции последнего на воздействия среды, сформировавшиеся в процессе эволюции. Приспособительный характер инстинктивных действий при этом также является результатом эволюционного процесса, так как постепенно закреплялись именно выгодные организму компоненты индивидуально-изменчивого поведения. С другой стороны, и сами инстинкты рассматривались Ламарком как изменчивые свойства животного. Тем самым взгляды Ламарка выгодно отличаются от встречаемых до наших дней воззрений на инстинкт как на воплощение неких сугубо спонтанных внутренних сил, обладающих изначально целесообразной направленностью действия. Что же касается индивидуально-изменчивых компонентов поведения животных, их «привычек», навыков, то Ламарк и здесь исходит из материалистических предпосылок, доказывая, что происхождение привычек обусловлено механическими причинами, лежащими вне организма. И хотя Ламарк был не прав, полагая, что сохраненные привычки видоизменяют организацию животного, но в его общем подходе к этой проблеме можно усмотреть правильное понимание ведущей роли функции по отношению к форме, поведения — по отношению к строению организма. Не давая здесь общей оценки эволюционному учению Ламарка, не касаясь недостатков и исторически обусловленных ошибок этого учения (изначальная целесообразность в природе, в частности в мире животных, гармоничность процесса развития, лишенного противоречий, и др.), необходимо, однако, подчеркнуть трудно переоценимую роль этого великого естествоиспытателя как основоположника материалистического изучения психической деятельности животных и развития психики в процессе эволюции. К. Ф. Рулье В середине XIX в. в России страстно и последовательно отстаивал исторический подход к изучению живой природы выдающийся ученый того времени, один из первых эволюционистов, профессор Московского университета К. Ф. Рулье. В те годы в естественных науках все большее распространение получали реакционные теории, и вопросы психики и поведения животных трактовались с идеалистических и метафизических позиций, преимущественно с точки зрения церковного учения. Психическая деятельность животных постулировалась как нечто раз и навсегда данное и неизменное; в их нравах и инстинктах усматривалась беспредельная, непостижимая мудрость Творца, все предусмотревшего и создавшего извечную гармонию в природе. В этих условиях засилья реакции Рулье решительно и обоснованно выступал против представлений о сверхъестественной природе инстинкта. Подчеркивая, что инстинкты необходимо изучать наравне с анатомией, физиологией и экологией животных, Рулье доказывал, что они являются естественной составной частью жизнедеятельности животных. Это особенно важно, если учесть, что в трудах того времени инстинкты, как и вообще поведение животных, трактовались в отрыве от среды обитания и образа жизни последних. В противоположность упомянутым тео- и телеологическим «объяснениям» Рулье усматривал первопричину происхождения психических качеств и особенностей животных во взаимодействии организма с условиями жизни. Зарождение и развитие инстинктов Рулье рассматривал как частный случай общей биологической закономерности, как результат материальных процессов, как продукт воздействия внешнего мира на организм. Инстинкт, по Рулье, — это сформировавшаяся на протяжении истории вида, выработанная условиями жизни наследственная реакция на определенные воздействия среды. Конкретные факторы происхождения инстинктов Рулье усматривал в изменчивости, наследственности и постепенном повышении уровня организации животного в ходе исторического развития. В индивидуальной жизни инстинкты могут изменяться в результате накопления опыта. Иллюстрируя изменчивость инстинктов, Рулье писал, в частности, что подобно тому, как меняются телесные качества животных, «подобно тому, как скот вырождается, как качества легавой собаки, неупражняемой, глохнут, так и потребность отлета для птиц, почему-либо долгое время не отлетавших, может утратиться: домашние гуси и утки сделались оседлыми, между тем как их дикие родичи — постоянно отлетные птицы. Лишь изредка отбившийся от дома домашний селезень во время сиденья его подруги на яйцах, начинает дичать, взлетывать и прибиваться к диким уткам; лишь изредка такой одичавший селезень отлетает осенью с своими родичами в теплую страну и следующею весной опять покажется на дворе, где вывелся».[18] Как перелетный инстинкт, так и нерестовое поведение рыб Рулье пытается проанализировать, опираясь на достижения естествознания его времени. Стремясь к полноценному естественноисторическому объяснению причины перелета птиц, он полагал, что птицы в свое время («когда на нашей планете начали слагаться климаты») начали выбирать себе наиболее благоприятные местности. Молодые птицы «и теперь отлетали бы от нас только в то время, когда голод, холод и нужда принудили бы их к этому, и полетели бы по тому направлению, по которому постепенно уменьшаются невыгодные для их жизни условия…».[19] Но старые птицы «помнят, как отлетали они прежде… знают, когда наступит срок для заблаговременного отлета… хотя неблагоприятные для них условия еще и не показывались».[20] Следовательно, в таком сложном явлении, как мифации птиц, Рулье усматривал сочетание и врожденных и благоприобретаемых компонентов, видового и индивидуального опыта. Интересно также отметить, какое значение Рулье придавал при этом передаче опыта от поколения к поколению. Он вообще считал, что у животных «обыкновение, сделавшееся в отце и матери естественною потребностью, отразится и на детях, как отражаются на них вообще привычки и наклонности родителей».[21] Оценивая зоопсихологические исследования Рулье, необходимо иметь в виду, что в его время (как, впрочем, и впоследствии) термином «инстинкт» часто прикрывали незнание истинных причин поведения животных. Сам Рулье говорил о том, «что инстинкт в большей части случаев — злоупотребляемое словцо, что за ним кроется вопрос, на который наука не нашла еще нужного объяснения…».[22] Поэтому Рулье стремился при каждом употреблении этого термина наполнить его конкретным содержанием, полученным в результате полевых исследований или экспериментов, делая при этом упор на выявление роли и взаимодействия факторов среды и физиологических процессов. Именно такой комплексный анализ поведения животных, стремление по возможности полнее выявить определяющие его экологические и исторические факторы и выдвинули труды Рулье на ведущее место среди естествоиспытателей середины прошлого века. Ч. Дарвин С победой эволюционного учения Ч. Дарвина в естествознании прочно утверждается мысль о единой закономерности развития в живой природе, о непрерывности органического мира. Сам Дарвин уделял большое внимание вопросам эволюции психической деятельности животных и человека. Им был написан фундаментальный труд «Выражение эмоций у человека и животных», а также ряд специальных работ по поведению животных. Для «Происхождения видов» Дарвин написал специальную главу «Инстинкт». О значении, которое Дарвин придавал изучению инстинктов, свидетельствует уже тот факт, что наличие их у человека и животных как общего свойства он рассматривал как одно из доказательств происхождения человека от животного предка. Дарвин воздерживался давать развернутое определение инстинкта, но все же указывал, что имеет при этом в виду такой акт животного, который выполняется им «без предварительного опыта или одинаково многими особями, без знания с их стороны цели, с которой он производится».[23] При этом он с полным основанием отмечал, «что ни одно из этих определений не является общим».[24] Ссылаясь на видного исследователя поведения животных первой половины XIX в. Фредерика Кювье (брата знаменитого зоолога и основателя палеонтологии позвоночных Жоржа Кювье), Дарвин сравнивает инстинкт с привычкой и указывает на ряд общих черт их проявления в противоположность сознанию. Но одновременно он предостерегает, что «было бы большой ошибкой думать, что значительное число инстинктов может зародиться из привычки одного поколения и быть наследственно передано последующим поколениям».[25] Происхождение инстинктов Дарвин объяснял преимущественным действием естественного отбора, закрепляющего даже совсем незначительно выгодные виду изменения в поведении животных и накапливающего эти изменения до образования новой формы инстинктивного поведения. Дарвин стремился показать, «что инстинкты изменчивы и что отбор может влиять на них и совершенствовать их».[26] Свою концепцию Дарвин сформулировал следующим образом: «Едва ли какой бы то ни было из сложных инстинктов может развиться под влиянием естественного отбора иначе, как путем медленного и постепенного накопления слабых, но полезных уклонений. Отсюда, как и в случае с органическими особенностями, мы должны находить в природе не постепенные переходы, путем которых развился каждый сложный инстинкт, — это можно было бы проследить только в ряде прямых предков каждого вида, — но должны найти некоторые указания на эту постепенность в боковых линиях потомков или, по крайней мере, мы должны доказать, что некоторая постепенность возможна, и это мы действительно можем доказать».[27] В подтверждение того, что происхождение даже весьма сложных форм инстинктивного поведения животных можно объяснить действием естественного отбора, Дарвин подвергает, в частности, обстоятельному анализу гнездовой паразитизм кукушки, «рабовладельческий инстинкт» некоторых видов муравьев и «самый удивительный из всех известных инстинктов — строительный инстинкт пчелы».[28] Действие «одного общего закона, обусловливающего развитие всех органических существ, именно размножения, изменения, переживания наиболее сильных и гибели слабых»,[29] Дарвин показывает и на многих других примерах поведения диких и домашних животных. Роль наследственности в процессе эволюции инстинктивного поведения он иллюстрирует такими фактами, как, например, обмазывание гнезда глиной у южноамериканских и британских дроздов или замуровывание самок в дуплах у птиц-носорогов Африки и Индии. Упражнениям и привычкам, т. е. индивидуальному научению, Дарвин, как уже отмечалось, не придавал сколько-нибудь существенного значения для исторического процесса становления инстинктивного поведения; он ссылался, в частности, на высокоразвитые инстинкты рабочих особей муравьев и пчел, неспособных к размножению и, следовательно, передаче накопленного опыта потомству. «Своеобразные привычки, присущие рабочим или бесплодным самкам, сколько бы времени они ни существовали, конечно, не могли бы воздействовать на самцов и плодущих самок, которые только и дают потомство»,[30] — писал Дарвин. «И меня удивляет, — продолжал он, — что до сих пор никто не воспользовался этим демонстративным примером бесполых насекомых против хорошо известного учения об унаследованных привычках, защищаемого Ламарком». Дарвин допускал возможность того, что лишь «в некоторых случаях свое влияние оказывают также и привычки и упражнение или неупражнение органа».[31] Дарвин был не прав, когда признавал единственным критерием развития природы лейбницкое «Natura non facit saltum» («Природа не делает скачков»), приложив этот принцип к инстинктам в равной мере, что и к строению организмов. Но, отстаивая идею взаимозависимости процессов в живой природе и доказывая их материальную сущность, Дарвин показал, что психическая деятельность животных подчиняется тем же естественноисторическим закономерностям, что и все другие проявления их жизнедеятельности. Очень важным является в этом отношении то, что Дарвин дал обоснованное естественнонаучное объяснение целесообразности инстинктов животных. Как и в отношении признаков строения организма, естественный отбор сохраняет, по Дарвину, полезные изменения врожденного поведения и упраздняет вредные. Эти изменения непосредственно связаны с морфологическими изменениями в нервной системе и в органах чувств, ибо конкретные формы поведения определяются особенностями строения нервной системы, которые передаются по наследству и подвергаются изменчивости, как и все прочие морфологические признаки. Таким образом, целесообразность инстинктов является результатом материального процесса — естественного отбора. Разумеется, это в корне противоречило теологическим воззрениям о сущности психического и его изначальной неизменности, в частности постулату о целесообразности инстинктов как проявлений божественной мудрости. Дарвин придерживался мнения, «что между развитием умственных способностей и инстинктов существует известное взаимодействие и что развитие последних предполагает некоторое унаследованное видоизменение мозга».[32] Прогресс умственных способностей, по Дарвину, обусловливался тем, что отдельные части мозга мало-помалу утратили способность отвечать на ощущения «определенным, однообразным, т. е. инстинктивным образом».[33] При этом Дарвин полагал, что инстинктивные компоненты тем больше господствуют у животных, чем ниже филогенетический ранг последних. Сегодня, спустя сто с лишним лет после этих высказываний Дарвина, мы не можем согласиться с таким противопоставлением основных категорий психической деятельности. Само разделение последней на «однообразно» выполняемые и изменчивые компоненты является условным, поскольку в каждом реальном поведенческом акте ригидные и лабильные элементы поведения выступают в едином комплексе. Соответственно на каждом филогенетическом уровне эти элементы, как будет еще показано, достигнут одинаковой степени развития. Современное понимание проблемы инстинкта и научения В проблеме инстинкта и научения большое место занимает вопрос о пластичности инстинктивного поведения. Этот вопрос является весьма важным для уяснения не только эволюции инстинктивного поведения, но и вообще всех вопросов, касающихся психической деятельности животных. Дарвин считал, что, по существу, одна пластичность инстинктов, проистекающая из изменчивости их врожденных морфологических основ и дающая «материал» для действия естественного отбора, является достаточной для эволюции инстинктивного поведения, а тем самым и поведения вообще. Впоследствии многие ученые посвятили свои усилия изучению того, насколько врожденное, видотипичное поведение является устойчивым или вариабельным, насколько инстинкты постоянны, ригидны или же изменчивы и могут модифицироваться. В результате сегодня мы знаем, что пластичность поведения животных является значительно более сложным явлением, чем это представлялось во времена Дарвина, ибо генетически фиксированы и передаются по наследству не отдельные готовые движения или их сочетания, а нормы реагирования, в пределах которых формируются двигательные реакции в онтогенезе. Глубокую разработку проблемы инстинкта и научения, как отмечалось, дал В. А. Вагнер, особенно в своем фундаментальном труде «Биологические основания сравнительной психологии» (1910–1913). Опираясь на большой фактический материал, полученный им в полевых наблюдениях и экспериментах и охватывающий как беспозвоночных, так и позвоночных, Вагнер пришел к выводу, что инстинктивные компоненты поведения животных возникли и развились под диктовку среды и под контролем естественного отбора и что их никак нельзя считать неизменными, стереотипными. Инстинктивное поведение, по Вагнеру, — это развивающаяся пластическая деятельность, изменяемая внешними воздействиями. Вариабельность инстинктивного поведения особенно убедительно была показана Вагнером на примерах конструктивной деятельности пауков и ласточек (рис. 4). Обстоятельный анализ этих фактов привел его к выводу, что лабильность инстинктивного поведения ограничена четкими видотипичными рамками, что стабильными в пределах вида являются не сами инстинктивные действия, а пределы амплитуд их изменчивости. Тем самым Вагнер предвосхитил одно из основных положений современной этологии. 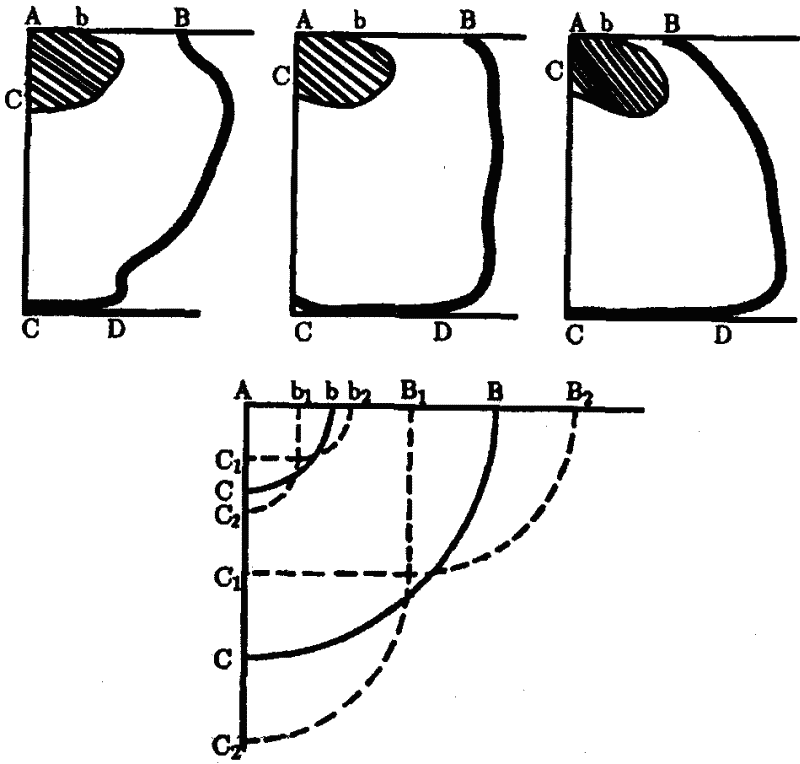 Рис. 4. Гнездостроение у городской ласточки (по Вагнеру). Верхний ряд — контуры разных форм ласточкиных гнезд. Постройки различаются, в частности, относительными размерами крыши (АВ) и дна (CD) гнезда (вид сбоку). Abc — леток. Внизу — схема Вагнера, изображающая вариабельность гнездостроительного поведения. Внутренняя дуга означает среднюю форму и величину летка, внешняя — гнезда. Пунктирные линии — отклонения по длине крыши и боковой стенке. Крайние точки (В1, B2, С1, С2; b1, b2 с1, c2) указывают на видотипичные пределы амплитуд изменчивости этих параметров при нормальном гнездостроении Рис. 4. Гнездостроение у городской ласточки (по Вагнеру). Верхний ряд — контуры разных форм ласточкиных гнезд. Постройки различаются, в частности, относительными размерами крыши (АВ) и дна (CD) гнезда (вид сбоку). Abc — леток. Внизу — схема Вагнера, изображающая вариабельность гнездостроительного поведения. Внутренняя дуга означает среднюю форму и величину летка, внешняя — гнезда. Пунктирные линии — отклонения по длине крыши и боковой стенке. Крайние точки (В1, B2, С1, С2; b1, b2 с1, c2) указывают на видотипичные пределы амплитуд изменчивости этих параметров при нормальном гнездостроении Впоследствии и другие советские ученые разрабатывали вопросы изменчивости инстинктивного поведения и его связи с процессами научения. Академик Л. А. Орбели проанализировал зависимость пластичности поведения животных от степени их зрелорождения (см. ч. II, гл. 3). Советский орнитолог А. Н. Промптов указывал на то, что инстинктивные действия животных (птиц и млекопитающих) всегда включают в себя неотъемлемые, очень трудно отчленяемые, но чрезвычайно существенные условнорефлекторные компоненты, формирующиеся в процессе онтогенеза. Именно эти компоненты, по Промптову, обусловливают пластичность инстинктивного поведения. С другой стороны, взаимодействие врожденных реакций, детерминированных видотипичными признаками строения, с приобретенными на их основе в течение индивидуальной жизни условными рефлексами дает в итоге видотипичные особенности, названные Промптовым «видовым стереотипом поведения». Е. В. Лукина иллюстрировала эти положения Промптова примерами по пластичности гнездостроительной деятельности воробьиных птиц. Так, молодые самки, гнездящиеся впервые в жизни, вьют гнезда, характерные для своего вида. Однако в необычных условиях этот стереотип заметно нарушается. Так, горихвостка и синица-пухляк, являющиеся дуплогнездниками, устраивают при отсутствии дуплистых деревьев свои гнезда под корнями, а серая мухоловка, вьющая гнезда в укрытиях (расщелинах пней, углубленных стволов, за отставшей корой и т. п.), может в случае необходимости устраивать их и на горизонтальных ветках или даже прямо на земле и т. д. Как мы видим, все это случаи модификации гнездостроительного инстинкта, конкретно — в отношении местоположения гнезда. Описано также немало примеров замены гнездостроительного материала: вместо травинок, мха, лишайника подчас используются такие искусственные материалы, как вата, упаковочные стружки, марля, веревки и т. п. Известны даже случаи, когда мухоловки-пеструшки сооружали свои гнезда в парках Москвы почти целиком из трамвайных билетов. Аналогичные данные были получены и в специальных опытах, в которых исследовалась пластичность инстинктивного поведения при замене яиц или птенцов (опыты Промптева, Лукиной, Скребицкого, Вилке). У насекомых пластичность строительной деятельности (сооружение чехликов у гусениц Psyche viciella) была в 60-х годах обстоятельно изучена в лаборатории польского зоопсихолога Р. И. Войтусяка (исследования К. Громыша). У другого насекомого, гусеницы Autispila stachjanella, там же изучалась пластичность инстинктивного поведения при устройстве ходов в листьях и коконах (исследования М. Берестынской-Вильчек). Ученые обнаружили большую адаптивную вариабельность инстинктивных действий, особенно при починке сооружений этих насекомых, причем в искусственных, экспериментальных условиях эти действия могут заметно отличаться от выполняемых в обычных, естественных условиях. Промптов был, безусловно, прав, когда подчеркивал значение слияния врожденных и приобретаемых компонентов во всех формах поведения. Вместе с тем его понимание пластичности инстинктов представляет собой шаг назад по сравнению с концепцией Вагнера, доказавшего, что врожденными являются не инстинктивные действия, а те рамки, в пределах которых эти действия могут выполняться в измененном виде в соответствии с данными условиями среды. Факты, приводимые Промптовым, лишь подтверждают это. Отказав инстинктивным компонентам поведения во всякой собственной вариабельности, Промптов считал, что пластичность обеспечивается лишь условнорефлекторными компонентами поведенческого акта. В действительности же мы имеем здесь дело с разными по своей величине и своему значению категориями изменчивости поведения. Во-первых, это изменчивость самих врожденных, т. е. инстинктивных, компонентов, которая проявляется в индивидуальной вариабельности видотипичного поведения (в пределах наследственно закрепленной нормы видотипичного реагирования). Примерами такой вариабельности могут служить приведенные Вагнером факты по гнездостроительной деятельности ласточек и упомянутые исследования инстинктивного поведения гусениц. Во-вторых, правда реже, в экстремальных условиях видотипичное, инстинктивное поведение может довольно сильно видоизменяться. Здесь проявляется роль индивидуального опыта, так как именно он обеспечивает более или менее выраженную приспособляемость к необычным, выходящим за рамки нормы внешним условиям. И, наконец, существуют различные индивидуально приобретаемые и поэтому максимально вариабельные формы поведения, в которых главенствующую роль играют уже разные формы научения, имеющие под собой, конечно, тоже инстинктивную основу и переплетающиеся с врожденными компонентами поведения. Принципиальное значение различий в изменчивости инстинктивного и благоприобретаемого поведения глубоко проанализировал академик А. Н. Северцов, основоположник эволюционной морфологии. В работах «Эволюция и психика» (1922) и «Главные направления эволюционного процесса» (1925) он показал, что у высших животных (млекопитающих) существуют два типа приспособления к изменениям окружающей среды: (1) изменение организации (строения и функций животных), совершающееся весьма медленно и позволяющее приспособиться лишь к очень медленно протекающим постепенным Изменениям среды, и (2) изменение поведения животных без изменения их организации на основе высокой пластичности ненаследственных, индивидуально приобретаемых форм поведения. В последнем случае возможно эффективное приспособление к быстрым изменениям среды именно благодаря изменению поведения. В этом случае наибольший успех будут иметь особи с более развитыми психическими способностями, «изобретатели» новых способов поведения, как метафорически выразился Северцов, — словом, животные, способные выработать наиболее гибкие, пластичные навыки и другие высшие формы индивидуального изменчивого поведения. Именно в этом плане Северцов рассматривает значение прогрессивного развития головного мозга в эволюции позвоночных. Что же касается инстинктивного поведения, то оно из-за своей малой изменчивости (ригидности) не может выполнять такую функцию. Но подобно изменениям строения тела животного изменения врожденного поведения могут служить приспособлением к медленным, постепенным изменениям среды, поскольку они требуют много времени для своего осуществления. Напомним, что это изменение врожденного поведения совершается в процессе эволюции на основе индивидуальной изменчивости инстинктивного поведения. Именно в этой индивидуальной лабильности видотипичных действий животных Дарвин искал источник зарождения и развития инстинктов. Северцов подчеркивал, что значение такого постепенного приспособления не менее важно, чем приспособление с помощью изменения индивидуально приобретаемого поведения. «Инстинкты, — писал он, — суть приспособления видовые, полезные для вида в такой же степени, как и те или другие морфологические признаки, и столь же постоянные».[34] При этом Северцов обращал внимание на то, что способность к научению, к установлению новых ассоциаций зависит от определенной наследственной высоты психической организации. Сами действия не являются при этом наследственными. В инстинктивном же поведении наследственно закреплено и то и другое. Сегодня мы можем в подтверждение этого тезиса Северцова сказать, что инстинктивное поведение животных определяется врожденной программой действий, реализуемой в ходе накопления индивидуального опыта. Подводя итог изложенному и учитывая современные знания о поведении животных, можно следующим образом охарактеризовать взаимосвязь и взаимозависимость между врожденными и приобретаемыми компонентами поведения и биологическое значение их специфической изменчивости. Постоянство, ригидность инстинктивных компонентов поведения необходимы для обеспечения сохранности и неуклонного выполнения наиболее жизненно важных функций независимо от случайных, преходящих условий среды, в которых может оказаться тот или иной представитель вида. Во врожденных компонентах поведения хранится итог всего эволюционного пути, пройденного видом. Это квинтэссенция видового опыта, самое ценное, что приобретено в ходе филогенеза для выживания особи и продолжения рода. И эти передаваемые от поколения к поколению обобщенные и генетически фиксированные программы действий не должны и не могут легко изменяться под влиянием случайных, несущественных и непостоянных внешних воздействий. В экстремальных же условиях еще имеются шансы выживания благодаря резервной пластичности инстинктивного поведения в виде модификации. В остальном реализация врожденной программы поведения в конкретных условиях индивидуального развития животного обеспечивается процессами научения, т. е. индивидуального приспособления врожденного, видотипичного поведения к частным условиям среды. Для этого необходима предельная гибкость поведения, но опять же возможность индивидуального приспособления без утраты существенного, накопленного в ходе эволюции вида, требует незыблемой основы в виде устойчивой инстинктивной диспозиции. Только она дает животному способность с выгодой для себя реагировать в любой ситуации. Глава 2 ИНСТИНКТИВНОЕ ПОВЕДЕНИЕ Инстинктивное поведение как основа жизнедеятельности животных Как было показано, инстинктивное поведение и научение не существуют в реальном поведении сами по себе, а только совместно, переплетаясь друг с другом в единый поведенческий акт. Однако это не означает, что инстинктивное поведение или научение представляет собой лишь условности, искусственно созданные для научного анализа. Условным является здесь лишь их разделение, сами же эти компоненты действительно существуют и функционируют как четко различимые процессы жизнедеятельности со своими специфическими качественными особенностями. В современных научных исследованиях все чаще избегают употребления понятия «инстинкт» ввиду многообразия и нечеткости его толкования. Понимаемое же как врожденное, наследственно фиксированное, видотипичное «инстинктивное поведение» складывается из инстинктивных действий или актов, которые в свою очередь состоят из отдельных инстинктивных движений (или поз, звуков и т. п.). Четкое разграничение этих терминов необходимо для этологического анализа обсуждаемой категории поведения животных. Когда мы говорим, что поведение является совокупностью функций внешних, «рабочих» органов животного организма, необходимо различать сами эти функции и их ориентацию во времени и пространстве. И то и другое происходит на инстинктивной основе. Научением же можно только менять ориентацию этих функций. Это означает, что никаким научением нельзя заставить функционировать органы животного иначе, чем это обусловлено их генетически фиксированным строением. Именно морфологические особенности определяют характер функционирования экзосоматических органов, т. е. инстинктивных движений. Нельзя вопреки поговорке научить зайца спички зажигать, так как у него нет соответствующих морфофункциональных предпосылок в строении его конечностей. Но можно научить зайца пользоваться своими конечностями естественным (инстинктивным) образом в нужный момент и в определенном направлении, т. е. ориентировать его инстинктивные движения во времени и пространстве путем научения (в данном случае — дрессировки). Сказанное не следует понимать в том смысле, что в поведении вообще первичным являются строение, структура, а вторичным — функция, движение. Наоборот, мы говорим о примате движения, функции, имея при этом в виду, что функция определяет форму. Биологическая обусловленность поведения не означает его морфологическую обусловленность; особенно в историческом, филогенетическом плане. В процессе эволюции, несомненно, поведение определяло формирование морфологических признаков, необходимых именно для более успешного выполнения самих поведенческих актов. Но когда мы говорим о конкретных инстинктивных движениях — результатах эволюционного процесса, то имеем в виду функции именно этих морфологических образований и то, что форма выполнения поведенческих функций обусловливается соответствующими морфологическими структурами. Конкретно это означает, что каждое животное может передвигаться или питаться только так, как это определяется специфическим строением его внешних органов, служащих для выполнения этих функций. Учитывая все это, можно сказать, что вся жизнедеятельность животного организма, проявляющаяся во внешней активности, зиждется на инстинктивных движениях и других инстинктивных реакциях (термических, электрических, изменения окраски, выделения секретов и т. п.). Они обеспечивают все жизненные функции организма, процессы обмена веществ, а тем самым существование особи и размножение. Вот почему мы говорим о примате инстинктивных движений и по отношению к нервной деятельности, сенсорике, психическому отражению, которые служат у животных лишь для осуществления этих движений, для их ориентации. Поэтому в эволюционном плане развитие психики являлось необходимым следствием (а затем уже — и предпосылкой) повышения уровня обмена веществ и двигательной активности. Инстинктивное поведение не исчерпывается, однако, самими функциями экзосоматических органов, а включает в себя и механизмы их регулировки и пространственно-временной ориентации. В этом отношении регулировка и ориентация, осуществляемые благоприобретенным путем, на основе научения, служат хотя и важным, но все же лишь дополнением к этим инстинктивным процессам. Внутренние факторы инстинктивного поведения Как уже упоминалось, проблема инстинкта и научения непосредственно связана с другой не менее важной проблемой — проблемой внутренних и внешних факторов, мотивации поведения. Долгое время считали, что инстинктивные действия определяются внутренними, притом загадочными, причинами, в то время как индивидуальное научение зависит от внешних стимулов. В таком виде представления об исключительном или хотя бы преобладающем значении внутренних или внешних факторов встречаются уже у античных мыслителей. При этом мистический, телеологический подход основывался на постулате изначальной целесообразности чисто внутренних, передаваемых от поколения к поколению факторов. Механистический же подход, получивший особое развитие со времен Декарта, признавал лишь внешние факторы движущими силами поведения. В ряде случаев и тот и другой взгляд непримиримо отстаивались вплоть до нашего века. Что же мы сегодня знаем о мотивации, о движущих силах инстинктивного поведения, а тем самым о поведении вообще? Обратимся сначала к тем внутренним факторам, которые дают первый толчок к любому поведенческому акту, не вдаваясь при этом в детали тех весьма сложных физиологических процессов, которые удалось в этой связи изучить за последние десятилетия. Внутренняя среда животного организма непрерывно обновляется, но, несмотря на непрекращающиеся процессы ассимиляции и диссимиляции, эта среда остается по своим физиологическим показателям постоянной. Стабильность внутренней среды — совершенно необходимое условие для жизнедеятельности организма. Только при этом условии могут осуществлятся необходимые биохимические и физиологические процессы. Любые, даже незначительные, отклонения от нормы воспринимаются системой интерорецепторов и приводят в действие физиологические механизмы саморегуляции, в результате чего эти нарушения устраняются. Такими механизмами саморегуляции советский физиолог академик П. К. Анохин считал сложные динамические структуры, функционирующие по принципу обратной связи (обратной афферентации) и обозначенные им как функциональные системы (рис. 5). 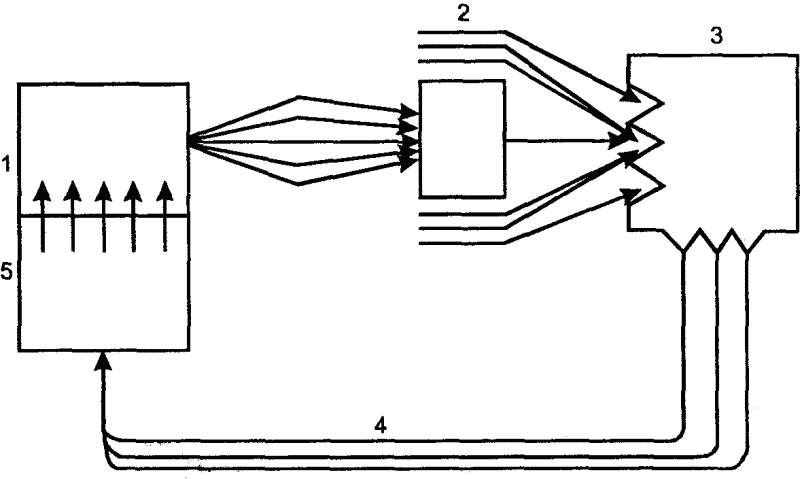 Рис. 5. Схема саморегуляции функциональной системы по Анохину: 1 — программа действия; 2 — результат; 3 — рецептор результата; 4 — обратная афферентация; 5 — афферентный синтез Рис. 5. Схема саморегуляции функциональной системы по Анохину: 1 — программа действия; 2 — результат; 3 — рецептор результата; 4 — обратная афферентация; 5 — афферентный синтез Итак, постоянство внутренней среды основано на самовосстанавливающейся уравновешенности внутренних процессов организма. Важная особенность этих процессов состоит в том, что они протекают в форме ритмов, которые также строятся на системах саморегулирования. Именно в сдвигах этих ритмов видный советский зоопсихолог В. М. Боровский еще в 30-е годы усматривал первичную мотивацию поведения. Выступая против идеалистического понимания инстинктивного поведения, он показал, что в мотивации этого поведения, т. е. в том, что принято называть побуждениями или влечениями, нет ничего сверхъестественного, оторванного от материального мира. Внутреннее побуждение, подчеркивал он, всегда является сдвигом корреляции физиологических ритмов в организме в сторону установления наиболее выгодной в данных условиях коррелированности ритмов всех физиологических процессов. В этом постоянном восстановлении внутреннего равновесия Боровский усматривал основу жизнеспособности организмов. Итак, первопричиной и основой мотивации поведения являются более или менее значительные и длительные отклонения от нормального уровня физиологических функций, нарушения внутренних ритмов, обеспечивающих жизнедеятельность организма. Эти сдвиги выражаются в появлении потребностей, на удовлетворение которых и направлено поведение. Первостепенное значение имеют для внутренней мотивации поведения ритмические процессы, происходящие в центральной нервной системе. Собственная ритмика ее стволовой части у позвоночных и брюшных нервных структур у беспозвоночных обеспечивает прежде всего ориентацию поведения во времени. Сейчас хорошо известны автономные, самовозбуждающиеся колебательные процессы («внутренние», или «биологические, часы»), которые регулируют общую ритмику жизнедеятельности организма. В отношении поведения это означает, что периодические колебания внешней активности животных, начало и окончание ритмично повторяющихся действий определяются ритмом «внутренних часов», синхронизированных с космическим временем. Существенные поправки или изменения вносятся в поведенческие ритмы многообразными биологически важными факторами среды, но общая «канва» инстинктивного поведения определяется самовозбуждающимися колебательными процессами с периодом, равным приблизительно суткам (околосуточный, «циркадный» ритм). В нормальных условиях этот ритм синхронизирован с изменениями в среде, определяемыми вращением Земли вокруг своей оси в течение суток. Однако и в искусственных условиях полной изоляции животного можно наблюдать обычную смену форм активности в те же сроки, что и в нормальных условиях. Это могут быть, например, изменения, связанные со сменой дня и ночи, хотя животное находится в эксперименте в условиях постоянного равномерного освещения. Помимо циркадных ритмов в поведении животных проявляются и более краткосрочные ритмы, многократно повторяющиеся в течение дня. Так, немецкий этолог В. Шлейдт установил, что клохтанье индюка вполне закономерно повторяется периодически даже в том случае, когда птица полностью изолирована от внешнего мира и даже лишена слуха. Конечно, в нормальных условиях протекание внутренних ритмов изменяется под влиянием внешних воздействий (слуховых, зрительных и прочих стимулов, метеорологических факторов и т. д.), а также зависит от общего физиологического состояния животного. «Внутренние часы» необходимы и для ориентации животных в пространстве. Хороший пример тому — ориентация птиц во время перелета. Руководствуясь, например, таким астрономическим ориентиром, как солнце, птицы должны учитывать его положение на небосводе в каждое данное время дня, что и происходит путем сопоставления воспринимаемой информации о положении солнца с фазами циркадного ритма. Уже упомянутый эксперимент Шлейдта показывает, что эндогенные процессы в нервной системе способны обусловливать выполнение определенных инстинктивных движений и при полном отсутствии адекватных внешних стимулов. Так, немецкий нейрофизиолог Э. Хольст обнаружил в стволовой части головного мозга курицы ряд зон, активация которых (в эксперименте — электрическим током) вызывает типичные инстинктивные движения различного функционального значения. При этом оказалось, что при раздражении одного и того же участка мозга, с усилением силы раздражения одно инстинктивное действие сменяется другим в естественной последовательности. Получилась цепь видотипичных движений, выполняемых курицей в определенной биологически значимой ситуации, например при виде приближающегося к ней наземного врага. При этом не только выполнение двигательных реакций, но и последовательность их появления точно так же соответствовала естественному поведению курицы: сперва лишь легкое беспокойство, потом подъем, усиленное беспокойство и, наконец, взлет (рис. 6). Учитывая, что все это происходило в отсутствие каких-либо адекватных внешних раздражителей, становится ясным, что на сугубо эндогенной основе могут выполняться не только отдельные инстинктивные движения, но и целые системы таких движений — инстинктивные действия. Конечно, в естественных условиях такие системы видотипичных, врожденных действий включаются воздействием внешних, экзогенных агентов, в нашем примере — действительным приближением врага, воспринимаемого экстероцепторами. В этом случае постепенное его приближение вызовет усиление раздражения соответствующих участков мозговых структур, что в эксперименте достигалось искусственно при помощи воздействий электрическим током. Таким образом, поведение в своей основе внутренне так же организовано, так же зиждется на системах биологической саморегуляции, так же закодировано в генетическом фонде вида, как и процессы, определяющие другие функции организма. В этом и проявляется единство всех форм жизнедеятельности животных.  Рис. 6. Последовательное проявление отдельных фаз защитного поведения курицы при продолжительном раздражении одного из участков стволовой части мозга. Длительность и интенсивность раздражения показаны темной полосой (опыты Хольста) Рис. 6. Последовательное проявление отдельных фаз защитного поведения курицы при продолжительном раздражении одного из участков стволовой части мозга. Длительность и интенсивность раздражения показаны темной полосой (опыты Хольста) Внешние факторы инстинктивного поведения Когда говорят об автономности внутренних факторов поведения, об их независимости от внешней среды, то необходимо помнить, что эта независимость является лишь относительной. Уже из приведенных опытов Хольста видно, что эндогенная активность не существует ни «сама по себе», ни «сама для себя»: значение этих спонтанных процессов в центральной нервной системе состоит в прениспозиции к возникновению жизненно важных ситуаций («в случае чего — все готово»). В результате животное способно по первому же сигналу незамедлительно и с максимальной пользой для себя реагировать на изменение в окружающей среде. Эта готовность обеспечивается тем, что соответствующие эндогенные системы периодически активируются как собственной ритмикой, так и внешними воздействиями (например, изменением продолжительности светового дня, повышением или уменьшением температуры и т. д.). Однако инстинктивные движения, по этологической концепции, заблокированы специальной системой «врожденных пусковых механизмов». Последние представляют собой совокупность нейросенсорных систем, обеспечивающих приуроченность поведенческих актов к биологически адекватным условиям среды (к «пусковой ситуации»). Как только животное оказывается в такой ситуации, соответствующий врожденный пусковой механизм обеспечивает распознавание, оценку и интеграцию специфических для данной инстинктивной реакции раздражителей, после чего наступает растормаживание, снятие «блокировки». Очевидно, одновременно происходит активация соответствующих нервных центров и снижение порогов их раздражимости. Характерной особенностью для врожденных пусковых механизмов является избирательность реагирования на внешние стимулы: они отзываются только на совершенно определенные комбинации раздражителей, которые только и могут вызвать биологически целесообразную реакцию. Иными словами, в сенсорной сфере существует некая «фильтрующая» функция, выражающаяся в специфической преадаптационной «готовности» воспринимать такие раздражители. Итак, благодаря врожденным пусковым механизмам внутренняя мотивация поведения получает «выход наружу», т. е. создается возможность без индивидуального опыта в биологически значимых ситуациях реагировать так, чтобы это способствовало сохранению особи и вида. Подводя итог сказанному, можно сказать, что под врожденным пусковым механизмом следует понимать совокупность нейросенсорных систем, обеспечивающих адекватность поведенческих актов по отношению к «пусковой ситуации»: настройку анализаторов на восприятие специфических раздражителей и распознавание последних, интеграцию соответствующих раздражений и растормаживание (или активацию) нервных центров, связанных с данным поведенческим актом. Внешние раздражители, составляющие в своей совокупности пусковую ситуацию, получили название «ключевых раздражителей», поскольку они подходят к своим врожденным пусковым механизмам, как ключ к замку. Ключевые раздражители являются такими признаками компонентов среды, на которые животные реагируют независимо от индивидуального опыта врожденными, видотипичными формами поведения, точнее, определенными инстинктивными движениями. В описанном поведении курицы это будут определенные общие признаки, свойственные всем ее наземным врагам. Кроме собственно растормаживающих ключевых раздражителей (их называют также «пусковыми раздражителями») различают также настраивающие ключевые раздражители, предварительно понижающие порог раздражимости нервных центров, причастных к данным действиям животного, а также направляющие ключевые раздражители, о которых пойдет речь при обсуждении таксисов. Общим свойством всех ключевых раздражителей является то, что это специфические элементарные признаки жизненно важных компонентов среды. Ключевыми раздражителями являются простые физические или химические признаки («просто» форма, размер, подвижность, цвет, запах и т. д.), или их пространственные отношения (взаиморасположение деталей, относительная величина и т. д.), или же векторы. Носителями этих признаков могут быть как другие животные, так и растения и объекты неживой природы. В последнем случае ключевые раздражители выполняют преимущественно направляющую функцию. Так, например, немецкий этолог Ф. Вальтер показал, что у детенышей антилоп ключевым стимулом, определяющим выбор места отдыха (лежа, неподвижно), является «что-то вертикальное» вне зависимости от того, что конкретно это за объект. Наилучшим образом изучены ключевые раздражители, носителями которых являются животные. Эти раздражители представляют особый интерес и потому, что являются первичными, генетически фиксированными элементами общения у животных. Так, в ставших уже классическими опытах голландского зоолога Н. Тинбергена, одного из основоположников современной этологии, с помощью макетов изучалась пищевая реакция («попрошайничество») птенцов серебристых чаек (клевание клюва родительской особи) и дроздов (вытягивание шеи и раскрытие клюва) при появлении родительской особи. В естественных условиях голодный птенец серебристой чайки клюет красное пятно на желтом клюве родителя, и тот в ответ отрыгивает пищу в рот птенцу. В опытах предъявлялась серия все более упрощаемых моделей — макетов. Первая модель точно воспроизводила внешний облик естественного носителя ключевых раздражителей, т. е. головы взрослой серебристой чайки с желтым клювом и красным пятном на нем. В последующих моделях путем проб постепенно исключались отдельные детали, и в результате макет становился все менее похожим на голову птицы (рис. 7). В конце концов остался лишь плоский красный предмет с продолговатым выступом. Но этот предмет оказался способным вызвать даже более сильную реакцию птенцов, чем исходная модель. Еще более эта реакция может быть усилена, если этот макет заменить тонкой белой палочкой, исчерченной поперечными темно-красными полосами. Ключевыми раздражителями в данном примере будут просто «красное» и «продолговатое». 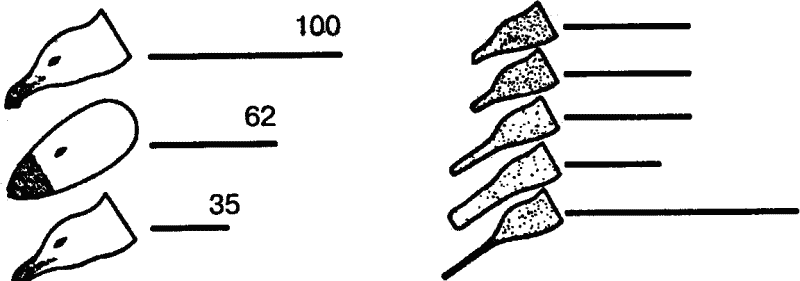 Рис. 7. Опыты Тинбергена с применением макетов головы серебристой чайки. Линиями обозначена интенсивность реакций птенцов на макет (число клеваний) Рис. 7. Опыты Тинбергена с применением макетов головы серебристой чайки. Линиями обозначена интенсивность реакций птенцов на макет (число клеваний) В опытах, проведенных Тинбергеном с десятидневными птенцами дроздов, выявилась другая категория ключевых раздражителей — взаиморасположение и относительная величина деталей объекта-носителя. В качестве макетов использовались плоские диски. Если показать птенцам такой круг, они будут тянуться к его верхней части (местонахождению головы птиц). Если же к большому кругу прибавить маленький, птенцы потянутся к нему. Если, наконец, прибавить к большому кругу два меньших круга, но разных размеров, то решающее значение приобретет относительная величина этих фигур. Размеры «головы» не должны приближаться к размерам «туловища»: при большом «туловище» птенцы потянутся к большему дополнительному кругу («голове») (рис. 8), при маленьком же—к меньшему. Интересные опыты проводил еще в середине 30-х годов Г. Л. Скребицкий совместно с Т. И. Бибиковой на подмосковном озере Киёво, изучая отношение чайки к ее яйцам. Яйца перекладывались из гнезда в гнездо, заменялись яйцами других видов, искусственными яйцами, а затем и различными посторонними предметами различной величины, формы и окраски. Оказалось, что чайки садились как на чужие, так и на деревянные, стеклянные, каменные, глиняные яйца различной величины и самой разнообразной окраски и начинали их «высиживать». То же самое наблюдалось, когда вместо яиц в гнездо подкладывались разноцветные шары, камешки или картофелины. По свидетельству экспериментаторов, «чайки, сидящие на таких предметах, представляли очень оригинальную картину, но в особенности необычайным становилось зрелище, когда согнанная с гнезда птица возвращалась к нему обратно и, прежде чем сесть, заботливо поправляла клювом разноцветные шары, камешки или картошку».[35] Если, однако, чайкам подкладывались предметы иной формы, например кубики или камни с неровными краями, поведение птиц заметно изменялось. Когда за край гнезда клалось по одному яйцу и инородному предмету, птицы вкатывали обратно в гнездо наряду с яйцами лишь округлые гладкие предметы, размеры которых соответствовали размерам яиц. В известных пределах не препятствовали этому существенные отклонения от нормы в весе (каменное яйцо весило в два с лишним раза больше, чем чаечье), материал, из которого был изготовлен предмет, и окраска.  Рис. 8. Опыты Тинбергена с птенцами дроздов Рис. 8. Опыты Тинбергена с птенцами дроздов Исследователи пришли к выводу, что положительная реакция чайки на яйцо определяется лишь несколькими его элементарными признаками: округлостью, отсутствием выступов, углублений или насечек (рис. 9). Именно эти признаки и выступали здесь как ключевые раздражители. 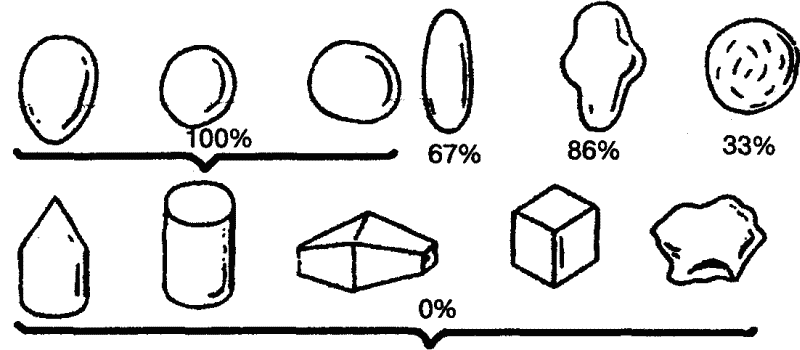 Рис. 9. Вкатывание в гнездо различных предметов в зависимости от их свойств у обыкновенной чайки (в процентах). Верхний ряд, слева направо: яйца (чаечьи, куриные, деревянные и каменные), деревянные шары, другие округлые предметы (картошка, камни) удлиненное глиняное яйцо с углублениями, глиняный шар с зарубками. Нижний ряд, слева направо: деревянный конус, деревянный цилиндр, деревянные и глиняные многоугольники, деревянные и глиняные кубики, угловатые камни (по Скребицкому и Бибиковой) Рис. 9. Вкатывание в гнездо различных предметов в зависимости от их свойств у обыкновенной чайки (в процентах). Верхний ряд, слева направо: яйца (чаечьи, куриные, деревянные и каменные), деревянные шары, другие округлые предметы (картошка, камни) удлиненное глиняное яйцо с углублениями, глиняный шар с зарубками. Нижний ряд, слева направо: деревянный конус, деревянный цилиндр, деревянные и глиняные многоугольники, деревянные и глиняные кубики, угловатые камни (по Скребицкому и Бибиковой) Важная особенность действия ключевых раздражителей заключается в том, что они подчиняются закону суммации: с увеличением их параметров пропорционально усиливается инстинктивная реакция животного. В экспериментальных условиях это может привести к так называемым «супероптимальным» реакциям, когда животное «преувеличенно», сильнее, чем в норме, реагирует на искусственный раздражитель, в котором «сгущены краски». Мы уже встречались с этим явлением при описании опыта с птенцами чайки, когда они сильнее реагировали на поперечно исчерченную красными полосками палочку, чем на настоящий клюв живой птицы. Суммация проистекает здесь из многократности красных меток и их большей контрастности. Эффект супероптимальной реакции может в условиях эксперимента привести даже к биологически абсурдному поведению животного. Если, например, предложить чайке два яйца разной величины, она закатит в гнездо более крупное. В итоге может возникнуть такое положение, что птица бросит свое яйцо, чтобы попытаться высиживать деревянный макет яйца гигантских размеров, обладающих супероптимальными признаками ключевого раздражителя. Как видно из приведенных примеров, ключевые стимулы действуют на поведение животного принудительно, заставляя его выполнять определенные инстинктивные движения, невзирая на возможно воспринимаемую животным общую ситуацию. Это объясняет многие, ранее казавшиеся загадочными моменты в поведении животных. Так, например, еще в начале нашего века один из авторов антропоморфических сочинений по психологии животных Т. Целл дал следующий ответ на вопрос о том, почему крупные хищники в нормальных условиях при встрече с человеком не нападают на него: например, лев не нападает не потому, что почитает человека, а потому, что не уверен в исходе схватки. «Есть ли у человека оружие, да какое оно, это оружие? — думает лев. — Да ну его, пойду-ка я лучше своей дорогой». Конечно, лев, как и другие крупные хищники, как правило, уклоняется от нападения на человека не потому, что руководствуется такими глубокомысленными рассуждениями. Разгадка «почтительного» отношения диких зверей к человеку, очевидно, кроется в следующем. Будучи сытым, хищник не реагирует и на присутствие животных, которыми обычно питается. У голодного же хищника преследование животных-жертв и нападение на них обусловливаются сочетанием рассмотренных выше внутренних факторов (первопричина — изменение уравновешенности внутренней среды организма в результате недостатка питательных веществ) с соответствующими внешними стимулами — ключевыми раздражителями, носителями которых являются естественные объекты питания, т. е. животные-жертвы, но не человек. Даже самый кровожадный хищник не волен нападать на кого угодно и когда угодно. Эти действия также ориентируются во времени и пространстве ключевыми раздражителями, как и все прочие поведенческие акты. Другими словами, дело не в том, «хочет» или «не хочет» животное поступить так или иначе. Если внутреннее состояние животного соответствует определенной внешней пусковой ситуации, то оно волей-неволей вынуждено вести себя так, как это диктует для данных условий генетически зафиксированный код видотипичного поведения. * * *Итак, в процессе эволюции возникают адаптации к более постоянным компонентам внешней среды, необходимым для удовлетворения потребностей, непрестанно возникающих в результате изменений внутренней среды организма. Нахождение (или избегание) важных для организма компонентов внешней среды осуществляется путем ориентации по типичным признакам этих компонентов — ключевым раздражителям. Результаты этой ориентации реализуются нейросенсорными системами (врожденными пусковыми механизмами), которые действуют рефлекторно и включают эндогенные, генетически фиксированные компоненты инстинктивного поведения. Таким образом, действуя «вовне», врожденные пусковые механизмы обеспечивают избирательную направленность внешней активности организма лишь на определенные сигнальные стимулы; действуя же «вовнутрь», они осуществляют оценку и отбор поступающей через рецепторы информации и ее реализацию для активации или понижения порогов раздражимости соответствующих нервных структур, для снятия «блокировки», растормаживания эндогенных нервных процессов, мотивирующих инстинктивные движения и действия. Таким образом осуществляется на врожденной основе корреляция внутренних потребностей организма с биологически существенными изменениями в окружающей его среде. Только в этой корреляции состоит все биологическое значение эндогенной мотивации поведения. Внутренние стимулы служат лишь для осуществления движения по отношению к среде, без которого организм — именно как саморегулирующаяся система — нежизнеспособен. И в этом, широком, смысле деятельность всей нервной системы в целом всегда рефлекторна. Равным образом даже наиболее как будто «далеко упрятанные» от внешней среды факторы поведения сами зависят от процессов обмена веществ. Эти процессы уже непосредственно связаны с окружающей организм средой. А так как организм активно регулирует, создает необходимые внешние предпосылки для нормального протекания обменных процессов как раз посредством поведения, то круг смыкается. И в этом отношении проявляется относительность независимости эндогенных автоматизмов, спонтанной активности нервной системы, способности к «самопрограммированию». Ясно, что все эти эндогенные процессы лишь постольку независимы от внешней среды, поскольку они лишь опосредованно связаны с ней. Структура инстинктивного поведения Поисковая и завершающая фазы поведенческого акта Выше говорилось о том, что ключевые раздражители действуют принудительно, что животное вынуждено в своем поведении всецело повиноваться пусковой ситуации. Но значит ли это, что у животных нет никакой возможности проявить собственную инициативу, произвести какой-то самостоятельный выбор? Отнюдь нет! Инициативное, избирательное отношение животного к среде проявляется прежде всего в активном поиске необходимых пусковых ситуаций и в избрании наиболее эффективных возможностей для выполнения поведенческих актов. Необходимо подчеркнуть, что речь идет о поиске именно раздражений, исходящих от биологически значимых объектов, а не самих этих объектов. Мы теперь уже знаем, что это ключевые раздражители с направляющей или пусковой функцией. Еще более полувека тому назад американский исследователь поведения животных У. Крэйг показал, что инстинктивные действия состоят из отдельных фаз. Прежде всего Крэйг выделил две фазы, которые в этологической литературе получили названия «поисковая» (или «подготовительная») и «завершающая». В ходе поисковой фазы животное отыскивает (отсюда и название фазы) те ключевые раздражители, точнее, их комбинации (т. е. пусковые ситуации), которые приведут его в итоге к завершающей фазе, в которой воплощается биологическое значение всего инстинктивного действия. Все промежуточные раздражители не составляют для животного самоцель и ценны лишь постольку, поскольку они приводят к восприятию ключевых раздражителей завершающего поведения. Только на завершающей фазе происходит собственно потребление животным жизненно необходимых ему элементов среды. Но сам поиск адекватных раздражителей является для животных такой же первейшей жизненной необходимостью, как и потребление элементов среды. Поисковая фаза всегда подразделяется на несколько этапов; однако, в завершающей же фазе, или вовсе не обнаруживаются такие подразделения, или же она состоит лишь из немногих строго последовательно выполняемых движений. Крэйг построил свою концепцию на данных, полученных им в результате изучения пищевого поведения животных. Приведем пример этой сферы поведения. Хищник, отправляющийся на охоту, вначале еще не знает, где находится его возможная добыча, поэтому первые его движения имеют характер ненаправленного поиска. В результате он раньше или позже попадает в сферу действия стимула, исходящего от животного-жертвы. Обнаружен первый ключевой раздражитель, который включает следующий этап — направленную ориентировку по дополнительным раздражителям, уточнение местонахождения животного-жертвы. Затем следует подкрадывание (или преследование), набрасывание (прыжок) и овладение добычей, ее умерщвление, иногда еще и перетаскивание туши в другое место, расчленение на отдельные куски и, наконец, захватывание кусков мяса зубами и их проглатывание. В этой цепи последовательно выполняемых действий и движений только последние два звена (собственно акт поедания) относятся к завершающей фазе описанного пищедобывательного поведения хищника, все остальные этапы составляют в своей совокупности поисковое (или подготовительное) поведение. Правда, внутри каждого такого этапа имеются свои подготовительные и завершающие фазы, которыми каждый этап и заканчивается. При этом подчас имеется несколько степеней соподчинения (по типу «матрешки»), так что в целом складывается весьма сложная структура деятельности. Аналогично обстоит дело и в других, казалось бы, значительно более простых сферах поведения, как, например, покой и сон. Животное вначале ищет место для отдыха или ночлега (деревья, укрытия, углубления в почве или просто определенные участки открытого пространства), затем устраивает (совершенствует) найденное место (копает, приминает растительность), иногда еще и чистится и только после этого укладывается (причем видотипичным образом!). Только укладывание составляет завершающую фазу, предшествующие же этапы — поисковую. К этим примерам можно было бы прибавить еще множество других из любой сферы поведения. Однако уже в приведенных можно усмотреть следующие глубокие различия между обеими фазами, определяющие их сущность. Поисковое поведение является пластичной фазой инстинктивного поведения. Она характеризуется ярко выраженной ориентировочно-исследовательской деятельностью животных и переплетением врожденных и приобретенных, основанных на индивидуальном опыте компонентов поведения. Именно к поисковому поведению относится все, что связано с пластичностью инстинктов, в частности с модификациями инстинктивного поведения. Завершающее поведение, наоборот, представляет собой ригидную фазу. Выполняемые в ней движения отличаются строгой последовательностью, стереотипностью и предопределены соответствующими макро- и микроморфологическими структурами. Приобретенные компоненты играют здесь несущественную роль или даже отсутствуют. Поэтому изменчивость ограничена индивидуальной (генетически фиксированной) вариабельностью. Сюда относится все, что говорилось о постоянстве, ригидности инстинктивного поведения и о принудительности действия ключевых раздражителей. Здесь практически все врожденное, генетически фиксированное. Что же касается движений, выполняемых в завершающей фазе, то это и есть собственно инстинктивные движения, или «врожденные двигательные координации», названные так австрийским ученым К. Лоренцем, одним из основателей современной этологии. Инстинктивные движения и таксисы Общая характеристика инстинктивных движений уже давалась выше. Также говорилось о том, что они являются «хранителями» самого ценного, жизненно необходимого, что накоплено видом в результате естественного отбора, и что именно этим определяется их независимость от случайных условий окружающей среды. Лисица, производящая инстинктивные движения закапывания мяса на каменном полу, ведет себя «бессмысленно». Но ведь не на каменном субстрате совершался филогенез лисиц, и было бы фатально для дальнейшего существования вида, если бы из-за случайного, временного пребывания особи в таких совершенно нетипичных для местообитания лисиц условиях исчезла бы столь полезная для этих животных форма поведения. Так что лучше произвести закапывающие движения при любых обстоятельствах, и «жесткая» врожденная программа поведения заставляет животное это сделать. Общая ориентация инстинктивных движений осуществляется таксисами, которые, по Лоренцу, всегда переплетаются с врожденными двигательными координациями и совместно с ними образуют единые инстинктивные реакции (или цепи из нескольких таких реакций). Как и инстинктивные движения, таксисы — это врожденные, генетически фиксированные реакции на определенные агенты среды. Но если инстинктивные движения возникают в ответ на пусковые раздражители, то таксисы отвечают на направляющие ключевые раздражители, которые неспособны обусловливать начало (или конец) какой-либо инстинктивной реакции, а лишь меняют вектор ее протекания. Таким образом, таксисы обеспечивают пространственную ориентацию двигательной активности животных в сторону благоприятных или жизненно необходимых условий среды (положительные таксисы) или же, наоборот, от биологически малоценных или опасных условий (отрицательные таксисы). У растений аналогичные реакции выражаются в изменениях направления роста (тропизмы). По характеру ориентирующих внешних стимулов таксисы подразделяются на фото-, хемо-, термо-, гео-, рео-, анемо-, гидротаксисы (реакции на свет, химические раздражители, температурные градиенты, силу тяжести, течение жидкости, поток воздуха, влажность среды) и т. д. На разных уровнях эволюционного развития таксисы имеют разную степень сложности и выполняют различные функции, о чем еще пойдет речь при обзоре эволюции психики. Сейчас важно подчеркнуть, что таксисы являются постоянными компонентами даже сложных форм поведения, причем высшие формы таксисов выступают в тесном сочетании с индивидуальным опытом животного. С направляющими ключевыми раздражителями мы уже встречались, когда описывалась ориентация птенцов по относительным оптическим стимулам. Само появление объекта (в эксперименте — диска, в естественных условиях — родительской особи) является пусковым раздражителем реакции «попрошайничества», взаиморасположение деталей этого объекта — направляющим ключевым раздражителем этой реакции, а пространственная ориентация птенцов по этому раздражителю — положительным фототаксисом. Точно так же красный цвет сам по себе является для птенца серебристой чайки пусковым ключевым раздражителем, обусловливающим его пищевую реакцию (наряду с другим пусковым раздражителем — появлением птицы, точнее, ее головы с клювом). Местонахождение же красного пятна на клюве направляет реакцию птенца биологически выгодным образом на основе положительного фототаксиса и служит, таким образом, направляющим ключевым раздражителем. Лоренц и Тинберген в 30-е годы совместно изучали взаимоотношения врожденных двигательных координации и таксисов на примере реакции вкатывания яиц в гнездо у серого гуся. Вид яйцеподобного предмета (нечто круглое, без выступов и т. д.), находящегося за пределами гнезда, служит у этой птицы таким же ключевым раздражителем для реакции вкатывания, как у сидящей на гнезде чайки в описанных опытах Скребицкого. Соответствующая врожденная двигательная координация представляет собой многократно повторяемое движение клюва к груди птицы, которое прекратится лишь тогда, когда предмет прикоснется к сидящей в гнезде птице. Если положить перед краем гнезда перпендикулярно к клюву цилиндр (гуси положительно реагировали и на такой предмет), то подобными инстинктивными движениями и ограничится все поведение птицы. Если же положить яйцо или его макет, то появляются дополнительные движения головы из стороны в сторону, придающие движению предмета правильное направление к гнезду. Ведь в отличие от цилиндра яйцо будет откатываться то влево, то вправо. Вид этих отклонений и служит направляющим раздражителем для таксисных боковых движений головы. Итак, таксисы могут у высших животных ориентировать инстинктивные движения не только всего организма, но и отдельных частей тела и органов. Из инстинктивных движений и таксисов складывается завершающая фаза каждого поведенческого акта. Вместе с тем они входят как составляющие части в поисковую фазу, которая, как уже говорилось, служит для поиска внешних пусковых ситуаций, позволяющих организму достигнуть завершающей фазы данного акта. Поисковая фаза отличается большой лабильностью и весьма сложной структурой. Инстинктивные движения завершают каждый промежуточный этап этой фазы, в результате конец каждого такого этапа также приобретает черты завершающего поведения. Таксисы дополняются в поисковой фазе ориентировочно-исследовательскими реакциями, непрерывно доставляющими организму информацию о состоянии, параметрах и изменениях компонентов окружающей среды, что позволяет ему произвести оценку последних в рамках общего поискового поведения. Благоприобретаемые компоненты инстинктивного акта Наряду с указанными в поисковой фазе любого инстинктивного акта всегда содержатся — в разной степени и в разных сочетаниях — и все элементы поведения, которые относятся к научению, не исключая и высших форм поведения, интеллектуального типа. Именно поэтому мы и считаем, что говорить об инстинктивных действиях (но не движениях!) — значит говорить о поведенческих актах вообще. В самом деле, все, чему животные способны научиться, направлено лишь к одному — по возможности к скорейшему и наиболее экономному достижению завершающего поведения. Нет в поведении животных ничего, что бы не завершалось этой конечной фазой — инстинктивными движениями или родственными им реакциями, т. е. врожденными двигательными координациями. И в этом отчетливо видно единство инстинктивного поведения и научения. Конечно, сказанное не относится в равной степени ко всем этапам поисковой фазы. Дело в том, что лабильность поведения неодинакова на разных этапах этой фазы и все больше уменьшается по мере приближения к завершающей фазе. Переход от одного этапа поискового поведения к другому означает активный поиск и нахождение животными ключевых раздражителей, смену одного этапа поискового поведения другим в строго закономерной последовательности. Это сопровождается ступенчатым сужением сферы активности животного: его поведение все более детерминируется специфическими для данного поведенческого акта сочетаниями раздражителей, становится все более направленным в сторону конечных, завершающих инстинктивных движений, суживаются возможности индивидуального видоизменения поведения животного, пока завершающая пусковая ситуация не сведет эти возможности практически к нулю. Так, например, ласточка, приступая к постройке гнезда, должна, прежде всего, найти место, где можно собирать гнездостроительный материал. Первоначально ненаправленный осмотр местности является первым этапом гнездостроительной деятельности. Скорость прохождения этого предельно лабильного этапа зависит, прежде всего, от индивидуально-изменчивых компонентов поведения, прежде всего от уже имеющегося индивидуального опыта. В зависимости от своих индивидуальных психических способностей каждая птица решает эту задачу по-своему, более или менее эффективно. Следующий этап поисковой фазы — поиск и сбор гнездостроительного материала на найденном подходящем месте. Здесь возможности индивидуального изменения видотипичного поведения уже сужены, но тем не менее индивидуальная сноровка продолжает играть немаловажную роль, Еще меньше амплитуда возможных индивидуальных отклонений на основе прежнего опыта на третьем этапе поискового поведения — транспортировке гнездостроительного материала к месту постройки гнезда. Здесь возможны лишь некоторые, не очень существенные вариации в скорости и траектории полета. В остальном поведение всех ласточек уже весьма стереотипное. И наконец, завершающая фаза — прикрепление частичек к субстрату — выполняется уже совершенно стереотипными инстинктивными движениями. Здесь только генетически обусловленная индивидуальная вариабельность видотипичного поведения дает уже известные нам различия в строении отдельных гнезд. Таким образом, постепенно уменьшается «свобода действия» животного, уменьшается амплитуда изменчивости его поведения по мере приближения к завершающей фазе в соответствии со все более ограниченными и специфическими условиями среды, сочетаниями раздражителей, причастных к данному поведению. Чем дальше от завершающей фазы и чем больше амплитуда изменчивости видотипичного поведения, тем больше возможностей включения в него элементов научения, индивидуального опыта и тем больше удельный вес этих элементов. Таким образом, индивидуальный опыт реализуется прежде всего на начальных этапах поискового повелении. И еще: чем выше психическое развитие, тем существеннее коррективы, вносимые в жесткое видотипичное поведение, но опять же главным образом на начальных этапах поисковой фазы. Все это относится, конечно, и к поисковому и завершающему поведению внутри каждого этапа поискового поведения. Сложность и разнообразие структуры инстинктивного поведения Двухфазная структура инстинктивных действий дана здесь лишь в виде весьма полной, упрощенной общей схемы. В реальной действительности чаще всего имеют место разные осложнения и видоизменения. Прежде всего необходимо иметь в виду, что поисковая фаза может происходить и под отрицательным знаком — в виде уклонения и избегания определенных агентов среды. Далее возможны сокращения поискового поведения, выпадения отдельных его этапов или даже инверсия. Иногда завершающие движения наступают так быстро, что поисковая фаза не успевает проявиться в полной мере. В других случаях поисковое поведение может свернуть со своего русла и привести к «чужому» завершающему поведению. Поисковое поведение может принять форму завершающего поведения и существовать наряду с истинной завершающей фазой. В таком случае внешне одинаковые действия будут иметь двоякую качественно различную мотивацию. Большой интерес представляют и различные случаи неполного протекания инстинктивного акта, когда действия животного не достигают завершающей фазы. У животных с наиболее высокоразвитой психикой промежуточные этапы поискового поведения, т. е. сам по себе поиск раздражителей, могут при этом — как исключение — стать самоцелью их поведения. Здесь мы встречаемся с инстинктивной основой наиболее сложных форм исследовательского поведения, входящих в фундамент интеллекта животных. Уже этот перечень показывает все разнообразие протекания инстинктивных действий; добавим еще, что практически никогда в одно и то же время не совершается лишь единственный инстинктивный акт, а имеет место сложное взаимодействие между несколькими одновременно происходящими действиями. Инстинктивное поведение и общение Все животные периодически вступают во внутривидовые контакты друг с другом. Прежде всего это относится к сфере размножения, где часто наблюдается более или менее тесный контакт между половыми партнерами. Кроме того, представители одного и того же вида часто скапливаются в местах с благоприятными условиями существования (обилие пищи, оптимальные физические параметры среды и пр.). В этих и подобных случаях происходит биологическое взаимодействие между животными организмами, на основе которого в процессе эволюции зародились явления общения. Ни любое контактирование между самцом и самкой, ни тем более скопление животных в благоприятных для них местах (зачастую с образованием колонии) не является проявлением общения. Последнее, как и связанное с ним групповое поведение, предполагает как непременное условие не только физическое или биологическое, но прежде всего психическое взаимодействие (обмен информацией) между особями, выражающееся в согласовании, интегрировании их действий. Как еще будет показано, это относится к животным, стоящим выше кольчатых червей и низших моллюсков. Дополняя сказанное, необходимо подчеркнуть, что об общении можно говорить лишь тогда, когда существуют особые формы поведения, специальной функцией которых является передача информации от одной особи к другой. Другими словами, общение, в научном значении этого термина, появляется лишь тогда, когда некоторые действия животного приобретают сигнальное значение. Немецкий этолог Г. Темброк, посвятивший много усилии изучению процессов общения и их эволюции, подчеркивает, что о явлениях общения и соответственно подлинных сообществах животных (стадах, стаях, семьях и т. д.) можно говорить лишь тогда, когда имеет место совместная жизнь, при которой несколько самостоятельных особей осуществляют вместе (во времени и пространстве) однородные формы поведения в более чем одной функциональной сфере. Условия такой совместной деятельности могут меняться, иногда она осуществляется при разделении функций между особями. Добавим еще, что если общение отсутствует у низших беспозвоночных и только в зачаточных формах появляется у некоторых их высших представителей, то, наоборот, оно присуще всем высшим животным (включая и высших беспозвоночных), и можно сказать, что в той или иной степени поведение высших животных в целом осуществляется всегда в условиях общения (хотя бы периодического). Как уже упоминалось, важнейшим элементом общения является обмен информацией — коммуникация. При этом информативное содержание коммуникативных действий (зоосемантика) может служить опознаванию (принадлежности особи к определенному виду, сообществу, полу и т. п.), сигнализировать о физиологическом состоянии животного (голоде, половом возбуждении и пр.) или же служить оповещению других особей об опасности, нахождении корма, места отдыха и т. д. По механизму действия (зоопрагматика) формы общения различаются каналами передачи информации (оптические, акустические, химические, тактильные и др.), но во всех случаях коммуникации животных представляют собой (в отличие от человека) закрытую систему, т. е. слагаются из ограниченного числа видотипичных сигналов, посылаемых «животным-экспедиентом» и адекватно воспринимаемых «животным-перцепиентом». Общение между животными невозможно без генетической фиксации способности как к адекватному восприятию (что обеспечивается таксисами), так и к передаче «кодированной» информации (что обеспечивается врожденными пусковыми механизмами). Наследственно закрепленные, инстинктивные действия, с помощью которых выполняется передача информации, могут приобретать самостоятельное значение. Здесь действуют те же закономерности, что и при других формах инстинктивного поведения, но носителями ключевых раздражителей являются в данном случае сородичи или животные других видов. Среди оптических форм общения важное место занимают выразительные позы и телодвижения, которые состоят в том, что животные весьма заметным образом показывают друг другу определенные части своего тела, зачастую несущие специфические сигнальные признаки (яркие узоры, придатки и т. п. структурные образования). Такая форма сигнализации получила название «демонстрационное поведение». В иных случаях сигнальную функцию выполняют особые движения (всего тела или отдельных его частей) без специального показа особых структурных образований, в других — максимальное увеличение объема или поверхности тела или хотя бы некоторых его участков (посредством его раздувания, расправления складок, взъерошивания перьев или волос и т. п.). Все эти движения всегда выполняются «подчеркнуто», нередко с «преувеличенной» интенсивностью (рис. 10). Но, как правило, у высших животных все движения имеют какое-то сигнальное значение, если они выполняются присутствии другой особи. Наиболее полноценная, четкая передача информации достигается, однако, тогда, когда появляются специальные двигательные элементы, выделившиеся из «обычных», «утилитарных» форм поведения, утративших в ходе эволюции свою первичную «рабочую», «механическую» функцию и приобретшие чисто сигнальное значение.   Рис. 10. Сигнальные позы и телодвижения у манящего краба (размахивание самцом клешней при «ухаживании» за самкой), рыбы (меченосца), полевого воробья и лошади (по Крейну, Хеменсу, Блуме и Хассенберг) Рис. 10. Сигнальные позы и телодвижения у манящего краба (размахивание самцом клешней при «ухаживании» за самкой), рыбы (меченосца), полевого воробья и лошади (по Крейну, Хеменсу, Блуме и Хассенберг) Первичные движения получили в этологии название «автохтонных» движений; вторичные, приобретшие новую, в данном случае сигнальную, функцию, — «аллохтонных». В таком случае сам рисунок стереотипно, у всех представителей данного вида одинаково выполняемого движения является условным выражением определенной биологической ситуации (биологически значимого изменения в среде или внутреннего состояния животного). Такие видотипичные стереотипные движения с четкой информативной функцией получили в этологии название «ритуализованных движений». Ритуализация характеризуется условностью выполняемых действий, которые служат лишь для передачи определенного, причем совершенно конкретного информативного содержания. Эти действия генетически жестко фиксированы, выполняются с максимальной стереотипностью, т. е., по существу, одинаково всеми особями данного вида, и поэтому относятся к типичным инстинктивным движениям. По этой причине все животные данного вида и в состоянии «правильно» понимать значение подобных оптических (или акустических) сигналов общения. Это типичный пример максимальной ригидности, консерватизма в поведении животных. Ритуализованные движения выполняются в строгой последовательности в форме более или менее сложных видотипичных «церемониалов» или «ритуалов», причем, как правило, в виде «диалога» двух животных. Чаще всего они встречаются в сферах размножения (брачные игры) и борьбы («мнимая борьба») и отображают внутреннее состояние особи и ее физические и психические качества. Наряду с врожденными существуют и благоприобретаемые формы общения, о которых пойдет речь позже. Все формы общения играют существенную роль в жизни высших беспозвоночных и позвоночных, обеспечивая согласованность действий особей. Без коммуникации с помощью различных химических, оптических, акустических, тактильных и других сигналов у этих животных невозможно даже сближение и контактирование самцов и самок, а, следовательно, и продолжение рода. Особое значение приобретает общение особей одного, реже разных видов в условиях их совместной, групповой жизни как основа группового поведения. Как уже говорилось, групповое поведение появляется (по меньшей мере в развитых формах) вместе с общением только у высших беспозвоночных и выражается в согласованных совместных действиях животных, живущих в сообществах. В отличие от скоплений животных, возникающих в результате идентичной положительной реакции на благоприятные внешние условия, сообщества характеризуются относительно постоянным составом его членов и определенной внутренней структурой, регулирующей отношения между ними. Эта структура нередко принимает форму иерархических систем соподчинения всех членов сообщества друг другу, чем обеспечивается сплоченность последнего и его эффективное функционирование как единого целого. Не меньшее значение имеет в этом отношении выделение вожаков, или «лидеров», регулирование отношений между молодыми и старыми животными, совершенствование ухода за потомством и т. д. Психический компонент инстинктивного поведения Психический компонент инстинктивного поведения, точнее, психическое отражение при инстинктивных действиях животных необходимо изучать на завершающей фазе поведенческого акта и в какой-то степени на непосредственно примыкающем к ней последнем этапе поисковой фазы, поскольку здесь элементы научения играют наименьшую роль. При этом надо сказать, что в свете современных данных и концепций этологии этот вопрос еще специально не изучался и поэтому приходится здесь ограничиваться лишь некоторыми общими соображениями. Мы видели, что по мере приближения к завершающей фазе движения животного становятся все более ограниченными, однообразными, стереотипными, и есть все основания полагать, что в такой же мере отражение окружающего мира становится все более бедным, ограниченным. Если на первых этапах поискового поведения животное в поиске ключевых раздражителей руководствуется многими разнообразными ориентирами в окружающей среде, то по мере сужения действия внешних агентов, направляющих поведение животного, ориентация все больше осуществляется только по направляющим ключевым раздражителям, а в завершающей фазе производится уже исключительно по ним. Вместе с тем мы теперь уже знаем, что как направляющие, так тем более и пусковые ключевые раздражители представляют собой элементарные физические и химические признаки, воспринимаемые животным «в отрыве» от объекта, которому они присущи. Следовательно, животное получает на завершающей фазе лишь весьма неполную, «однобокую» информацию о некоторых — причем несущественных — внешних признаках объектов инстинктивных действий. Правда, эти признаки являются постоянными и в этом отношении характерными для этих объектов. Однако, что для нас сейчас важно, животное, реагируя на эти признаки, в сущности, не получает сведений о самом объекте (неодушевленном предмете или другом живом существе): признаки — ключевые раздражители — являются только ориентирами, направляющими действия животного на этот объект, носитель этих ориентиров. Это с совершенной очевидностью вытекает из приведенных выше экспериментов, в которых подлинные объекты инстинктивных действий заменялись весьма абстрактными макетами. Вспомним, что в этих условиях удалось получить даже сверхоптимальные «преувеличенные» инстинктивные реакции. Самец колюшки в завершающей фазе борьбы за гнездовой участок не видит разницы между живой рыбой-соперником и куском жести или пластмассы. Он видит только «красное», и этого ему в данной ситуации вполне достаточно, чтобы среагировать на это «красное», которое в данном случае, кстати, выступает одновременно как направляющий и пусковой ключевой раздражитель. Следовательно, здесь налицо первично генерализованная реакция на общий элементарный признак (точнее, на совокупность нескольких таких признаков, поскольку в природных условиях действуют не одно только «красное», но и некоторые другие ключевые раздражители). Этот признак воспринимается недифференцированно, как простое ощущение (точнее, воспринимаются суммы ощущений). Об этом свидетельствует отсутствие реакций на форму объекта — носителя признака, да и вообще на объект (предмет или рыбу) как таковой. Ясно, что здесь обнаруживается очень бедное, крайне поверхностное и ограниченное отражение окружающего мира, низшая форма психического отражения. Неинтересно, что такое отражение, свойственное низшим представителям животного мира, в полной мере сохранилось и у высших животных — позвоночных и продолжает у них играть главную роль в поддержании жизнедеятельности особи и продолжении рода. О том, как это понимать, уже говорилось выше. Эволюция животной жизни могла идти только по пути отбора как можно более простых, но и наиболее постоянных ориентиров (или, что не меняет существа дела, создания таких специфических ориентиров, когда речь идет о животных одного вида, являющихся носителями ключевых признаков), руководствуясь которыми организмы могли бы успешно, не отвлекаясь другими, случайными, изменчивыми признаками, осуществлять наиважнейшие жизненные функции. Правда, как явствует, например, из описанных опытов с птенцами дроздов, у высших животных ключевые стимулы представлены и более сложными по своей структуре типами. В указанном примере имеет место реакция на конфигурацию и на пространственные соотношения деталей объектов инстинктивных движений, что уже соответствует элементарным восприятиям. Это, очевидно, одно из важных проявлений прогрессивного развития инстинктивного поведения в ходе эволюции. Однако, как показывают те же эксперименты, и здесь нет подлинного предметного восприятия, ибо иначе птенцы не могли бы спутать свою мать с картонным диском. Примитивность психического отражения на завершающей фазе инстинктивных действий является следствием бедности самой моторики в этой фазе. Как мы знаем, двигательная активность, направленная на окружающую среду, является источником познания этой среды. Однако столь стереотипные движения со столь ограниченной, специальной функцией, какими являются инстинктивные движения, врожденные двигательные координации, не могут служить сколько-нибудь пригодной основой для познания окружающего мира. Врожденные двигательные координации (во всяком случае в их типичной форме) и не выполняют эту функцию, а ключевые раздражители, воспринимаемые на завершающей фазе (да и на отдельных этапах поискового поведения), не познаются, а узнаются врожденным образом благодаря наличию врожденных пусковых механизмов, чтобы затем использоваться как ориентиры или — и того меньше — как «кнопки» пуска инстинктивных реакций. Здесь больше ничего нет, кроме, вероятно, положительной или отрицательной эмоциональной окраски ощущений от воспринимаемых стимулов и собственных движений. Познавательные же функции и вообще все богатство психического отражения приурочены к начальным этапам поискового поведения, где в полной мере действуют процессы научения. Глава 3 НАУЧЕНИЕ Общая характеристика процесса научения Анализ инстинктивного поведения приводит к выводу, что все богатство и разнообразие полноценного психического отражения связаны с научением, накоплением индивидуального опыта. Поэтому эта категория поведения будет постоянно в центре нашего внимания и мы еще не раз обратимся к разным ее аспектам, познакомимся с разными формами научения. Здесь же мы рассмотрим лишь некоторые общие положения. При изучении процессов научения необходимо исходить из того, что формирование поведения представляет собой процесс конкретного воплощения в жизни особи опыта вида, накопленного и закрепленного в процессе эволюции. Именно этим, как уже отмечалось, определяется переплетение врожденных и индивидуально приобретаемых компонентов поведения. Видовой опыт передается из поколения в поколение в генетически фиксированном виде в форме врожденных, инстинктивных компонентов поведения, поэтому «закодированное» таким образом поведение всегда является видотипичным. Но это не означает, что справедливо и обратное, т. е. что каждая видотипичная форма поведения по своему происхождению обязательно является врожденной, инстинктивной. Существуют такие формы научения, которые внешне очень напоминают инстинктивное поведение, но тем не менее представляют собой не что иное, как результат накопления индивидуального опыта, правда, в жестких видотипичных рамках. Это прежде всего формы так называемого облигатного научения, которым, по Г. Темброку, обозначается индивидуальный опыт, необходимый для выживания всем представителям данного вида независимо от частных условий жизни особи. Относящиеся сюда формы научения будут рассмотрены при обсуждении онтогенеза поведения. В противоположность облигатному факультативное научение, согласно Темброку, включает в себя все формы сугубо индивидуального приспособления к особенностям тех конкретных условий, в которых живет данная особь. Понятно, что эти условия не могут быть одинаковыми для всех представителей данного вида. Служа, таким образом, максимальной конкретизации видового поведения в частных условиях среды обитания вида, факультативное научение является наиболее гибким, лабильным компонентом поведения животных. О ряде важных аспектов факультативного научения также будет говориться более подробно в связи с онтогенезом поведения (ч. II, гл. 3). Но следует сразу оговорить, что лабильность факультативного научения неодинакова в разных формах его проявления. Конкретизация видового опыта путем вплетения в инстинктивное поведение элементов индивидуального опыта совершается на всех, даже ригидных, этапах поведенческого акта, хотя, как мы уже знаем, в разной степени. Так, английский этолог Рахайнд указывает на модификацию инстинктивного поведения процессами научения путем изменения сочетания раздражителей, обусловливающих определенный инстинктивный акт, выделения (или усиления) определенных направляющих ключевых раздражителей и т. п. При этом существенно, что модификации могут иметь место как в сенсорной, так и в эффекторной сфере, хотя чаще всего охватывают обе сферы одновременно. В эффекторной сфере включение элементов научения в инстинктивные действия происходит чаще всего в виде перекомбинации врожденных двигательных элементов, но могут возникнуть и новые двигательные координации. Особенно часто это происходит на более ранних этапах онтогенеза. Примером возникновения таких новых двигательных координации может служить подражательное пение птиц. У высших животных, как мы еще увидим, вновь выученные движения эффекторов, особенно конечностей, играют особенно большую роль в познавательной деятельности, в интеллектуальном поведении. Что же касается «встраивания» индивидуально-изменчивых компонентов в инстинктивные действия в сенсорной сфере, то это существенно расширяет возможности ориентации животного благодаря овладению новыми сигналами. При этом в отличие от ключевых раздражителей, которые являются для животного изначально биологически значимыми, раздражители, которыми животное руководствуется при научении, первоначально являются для него индифферентными (или, точнее, почти таковыми). Лишь по мере того как животное запоминает их в ходе накопления индивидуального опыта, они приобретают для него сигнальное значение. Таким образом, процесс научения характеризуется избирательным вычленением некоторых «биологически нейтральных» свойств компонентов среды, в результате чего эти свойства приобретают биологическую значимость. Сейчас мы уже знаем, что этот процесс не сводится просто к образованию условных рефлексов. Дело обстоит значительно сложнее, прежде всего благодаря поистине активному избирательному отношению организма к элементам окружающей среды. Основой этому служат многообразные динамические процессы в центральной нервной системе, преимущественно в высших ее отделах, благодаря которым осуществляется афферентный синтез раздражений, обусловленные внешними и внутренними факторами. Эти раздражения сопоставляются с ранее воспринятой информацией, хранимой в памяти. В результате формируется готовность к выполнению тех или иных ответных действий, что и проявляется в инициативном, избирательном характере поведения животного по отношению к компонентам среды. Следующий затем анализ результатов действий животного (по принципу обратной связи) служит основой для нового афферентного синтеза. В результате в дополнение к врожденным, видовым программам поведения в центральной нервной системе постоянно формируются новые, благоприобретаемые, индивидуальные программы, на которых зиждутся процессы научения. Физиологические механизмы этого непрерывного программирования поведения были особенно детально проанализированы П. К. Анохиным и получили свое отражение в его концепции «акцептора результатов действия». Весьма важным является здесь то, что животное не обречено на пассивное «слепое повиновение» воздействиям среды, а само строит свои отношения с компонентами среды и обладает при этом известной «свободой действия», точнее, свободой поиска и выбора. Итак, формирование эффективных программ предстоящих действий является результатом комплексных процессов сопоставления и оценки внутренних и внешних раздражений, видового и индивидуального опыта, регистрации параметров совершаемых действий и проверки их результатов. Это и составляет основу научения. Как мы уже знаем, реализация видового опыта в индивидуальном поведении в наибольшей степени нуждается в процессах научения на начальных этапах поискового поведения. Ведь именно на этих первых этапах каждого поведенческого акта поведение животного является наиболее неопределенным, именно здесь от него требуется максимальная индивидуальная ориентация среди разнородных и переменчивых компонентов среды, и особенно важен быстрый выбор наиболее эффективных способов действия, чтобы быстрее и наилучшим образом достигнуть завершающей фазы поведенческого акта. При этом животное может положиться только на собственный опыт, ибо реакции на единичные, случайные признаки каждой конкретной ситуации, в которой может оказаться любой представитель вида, конечно, не могли быть запрограммированы в процессе эволюции. Но поскольку без включения благоприобретаемых элементов в инстинктивное поведение последнее неосуществимо, само это включение также наследственно закреплено, как и инстинктивные компоненты поведения. Другими словами, диапазон научения также является строго видотипичным. Представитель любого вида не может научиться «чему угодно», а только тому, что способствует продвижению к завершающим фазам видотипичных поведенческих актов. Существуют, следовательно, видотипичные, генетически фиксированные «лимиты» способности к научению. Рамки диспозиции к научению не только соответствуют реальным условиям жизни, но у высших животных они подчас и шире, чем того требуют эти условия. Поэтому высшие животные обладают значительными резервами лабильности поведения, потенциальными возможностями индивидуального приспособления к экстремальным ситуациям. У низших животных эти возможности резко сужены, как и вообще способность к научению не достигает у них столь высокого уровня, как у высших животных. Поэтому широта диапазонов способности к научению, наличие указанных резервных возможностей являются показателями высоты психического уровня животных. Чем шире эти диапазоны и чем больше потенциальных возможностей к научению сверх обычной нормы, к накоплению и применению индивидуального опыта в необычных условиях, тем больше и лучше может быть скорректировано инстинктивное поведение, тем более лабильной будет поисковая фаза, тем более пластичным будет все поведение животного и тем выше его общий психический уровень. Это процесс диалектического взаимодействия, при котором причина и следствие постоянно меняются местами: усложнение в процессе эволюции инстинктивного поведения требует расширения диапазона способности к научению; усиленное же включение элементов научения в инстинктивное поведение делает в результате последнее более пластичным, т. е. приводит к качественному обогащению этого поведения, поднимает его на более высокую ступень. В результате поведение эволюционирует как единое целое. Эти эволюционные преобразования охватывают как содержание врожденных программ поведения, так и возможности их обогащения индивидуальным опытом, научением. На низших этапах эволюции животного мира эти возможности ограничиваются явлениями привыкания, тренировки и суммационными процессами в сенсорной сфере (см. ч. III). Появляющиеся у низших беспозвоночных временные связи еще весьма неустойчивы, затем они становятся все более прочными, достигая полного развития в разного рода навыках. При привыкании постепенно исчезают реакции на определенное многократно повторяющееся раздражение, не сопровождающееся биологически значимым воздействием на животное. Противоположным явлением можно считать тренировку, при которой инстинктивное действие совершенствуется путем накопления индивидуального опыта. Разница в поведении между низшими и высшими животными состоит не в том, что более примитивные формы поведения сменяются более сложными, а в том, что к первым добавляются вторые, существенно обогащая и изменяя их, в результате чего элементарные формы поведения приобретают значительную сложность и становятся более вариабельным. Так привыкание, с элементарной формой которого мы еще познакомимся на примере поведения простейших, выступает у млекопитающих в весьма сложных проявлениях. Например, как показали опыты, проведенные Хайндом совместно со Скаурсом, на мышах, реакции этих животных на многократно повторяемые, но ничем не подкрепленные акустические стимулы ослабевают с неодинаковой быстротой. Но эти различия в привыкании обусловливаются не только различиями в интенсивности стимулов (как это имеет место у низших животных), но прежде всего вариабельностью самого процесса привыкания. Основным качественно новым компонентом научения, дополняющим и обогащающим первичные примитивные формы поведения, является навык, который, однако, на разных эволюционных уровнях тоже проявляется в существенно различающихся формах. Навык Основные особенности навыков Навык — центральная, важнейшая форма факультативного научения. Иногда термин «навык» употребляется для обозначения научения вообще. Однако, как подчеркивает А. Н. Леонтьев, нельзя называть навыками любые связи, возникающие в индивидуальном опыте, ибо при таком расширенном понимании понятие навыка становится весьма расплывчатым, а тем самым непригодным для строго научного анализа. Уже указывалось на то, что способность к выработке навыков свойственна не всем животным, а проявляется лишь на определенном уровне филогенеза. В результате формирования навыка применяется врожденная двигательная координация (обычного поведения) в новой сигнальной ситуации или возникает новая (приобретенная) двигательная координация. В последнем случае появляются новые, генетически не фиксированные движения, т. е. животное научается что-то делать по-новому. Но в любом случае решающее значение имеет успешность выполняемых движений и их подкрепление положительным результатом. При этом, по существу, неважно, строится ли научение на информации, полученной путем собственного активного поиска раздражителей, либо в ходе общения с другими животными, коммуникаций, куда относятся также случаи подражания или обучения (человеком — животного, взрослой особью — детеныша и т. п.). Другой важнейший признак навыка, также связанный с необходимостью подкрепления, заключается в том, что он формируется в результате упражнения и нуждается для своего дальнейшего сохранения в тренировке. При тренировке навыки совершенствуются, при отсутствии же ее угасают, разрушаются. Навыки интенсивно изучаются у животных с помощью разнообразных специальных методов: «лабиринта», «проблемного ящика» (или клетки), «обходного пути» и др. При всех этих методах животное ставится в условия выбора сигналов или способов действия при решении определенной задачи. Уже описанный выше метод «лабиринта» был введен в практику экспериментального исследования в 1901 г. американским психологом В. С. Смоллом. В настоящее время применяются лабиринты самой различной конструкции (см. рис. 1), но задача, которую приходится решать подопытному животному, всегда одна и та же — как можно быстрее и без захода в тупики достигнуть места подкрепления (чаще всего пищевого). Большинство опытов проводилось с крысами, которые легко справляются с такими задачами. Практически не существует лабиринта, в котором крыса не сумела бы сориентироваться. Особенно интенсивно пользовались лабиринтами, как и проблемными ящиками и клетками (также уже описанными), бихевиористы. Проблемный ящик был усовершенствован и снабжен автоматическими устройствами американским психологом Б. Ф. Скиннером. В качестве подопытных животных при этом также использовались крысы. В лабиринте животное решает задачу как бы «вслепую», так как реакция (пробежка) вырабатывается без непосредственного контакта с обусловливающим ее стимулом (пищей, гнездом), и более того, животное в начале опыта еще ничего не знает о наличии целевого объекта, а лишь случайно обнаруживает его впоследствии в результате ориентировочно-исследовательских действий. Запоминание этого объекта и пути к нему и служат основой формирования навыка. Если впоследствии животное многократно пробежит это расстояние одним и тем же выученным кратчайшим путем, то этот навык становится стереотипным, автоматизированным. Пластичность поведения в этом случае невелика, а при сильно выраженной стереотипности выученных движений последние подчас приближаются к инстинктивным двигательным стереотипам. Стереотипность вообще характерна для примитивных навыков, при которых формируются ригидные, автоматизированные двигательные реакции. Значительная пластичность проявляется при таких навыках лишь па первых этапах образования этих навыков. (С навыками высшего порядка, отличающимися большой пластичностью, мы познакомимся позже.) Выработка навыков по методу Скиннера получила название «оперантное», или «инструментальное», обусловливание. Животное должно при решении таких задач проявить двигательную инициативу, самостоятельно «изобрести» способ действия, оперирования. Животному здесь не «навязываются» экспериментатором определенные движения, как это имеет место при выработке «классических» условных рефлексов по методу Павлова. При обучении крысы в скиннеровском «проблемном ящике» временная связь образуется после ряда случайных движений (нажатий на планочку), подкрепляемых появлением пищи в кормушке. В более сложно устроенных установках крысам предоставляется даже возможность выбора между двумя способами действия, приводящими к разным результатам. Так, например, можно крысе дать возможность «по своему усмотрению» регулировать температуру в «проблемном» ящике, включая то обогревающий рефлектор, то охлаждающий вентилятор в соответствии со своими потребностями. При каждом нажатии на рычаг попеременно включается рефлектор и выключается вентилятор или наоборот (рис. 11). При «классическом», павловском обусловливании наблюдается «респондентное» поведение, т. е. ответ следует за стимулом, а в результате образования условно-рефлекторной связи и подкрепление (безусловный раздражитель) связывается со стимулом. При оперантном же обусловливании вначале производится движение (ответ), сопровождаемое подкреплением без условного раздражителя. Но как и при «классической» выработке условных рефлексов, адекватная реакция животного (в данном случае двигательная) подкрепляется здесь полезным для животного результатом. При этом потребность в пище или создании оптимальных температурных условий побуждает крысу реагировать на рычаг выученным образом, точнее, ориентировать свое поведение в соответствии с восприятием рычага. Это восприятие действует как условное раздражение постольку, поскольку действие рычагом приводит к биологически значимому результату — пищевому или температурному подкреплению. Вне этой временной связи рычаг не имеет для крысы никакого значения.  Рис. 11. Крыса в скиннеровском «проблемном ящике» (объяснения см. в тексте) Рис. 11. Крыса в скиннеровском «проблемном ящике» (объяснения см. в тексте) Тот факт, что при павловском обусловливании действует сначала условный агент (условный раздражитель, например звук метронома или вспышка лампочки), а уже затем безусловный (например, вкусовое ощущение в результате пищевого подкрепления), не следует переоценивать, так как и условный рефлекс может быть выработан лишь при наличии внутренней готовности животного, т. е. соответствующей потребности. У вполне сытого животного невозможно выработать условный рефлекс на пищу. Выше уже говорилось о роли первичной (инстинктивной) мотивации в поведении животных. По существу, оперантное обусловливание по Скиннеру в сравнении с «классической» павловской постановкой эксперимента охватывает лишь упрощенный, укороченный поведенческий акт: вне внимания оставляется первая фаза — фаза ориентации животного во времени и пространстве. При выработке «классического» условного рефлекса животное должно сперва по внешним ориентирам научиться тому, когда, при каких сопутствующих внешних условиях следует произвести соответствующее движение. Вот эта первичная ориентация животного во времени по внешним сопутствующим признакам не учитывается при упомянутой постановке опытов по методу Скиннера. Равным образом не учитывается и пространственная ориентация, т. е. как животное находит рычаг, как научается пользоваться им, — словом, как ориентирует свои движения в пространстве. А ведь это является самым важным для подлинного психологического анализа поведения животного. Вместо такого анализа структуры деятельности при формировании двигательного навыка бихевиористами учитываются лишь временные параметры этого процесса (например, число нажатий за определенное время) без соответствующего качественного анализа. В противоположность этому «классическая» павловская методика уже по самому существу своему принципиально верно моделирует естественные условия формирования поведения животного, так как направлена на анализ первичной ориентации животного по признакам компонентов среды, по которым животному необходимо ориентироваться в самом начале любого поведенческого акта. Преимущества такого подхода становятся ясными, если вспомнить, что говорилось выше о первом этапе поискового поведения. Конечно, как отмечалось, мы сегодня уже не можем при изучении процессов научения, в частности навыков, ограничиваться «классическими» условными рефлексами. И разумеется, «проблемные ящики», лабиринты и тому подобные установки совершенно необходимы для изучения формирования навыков в разных условиях, для выявления способностей животных к решению задач разной степени сложности. Важно только наряду с количественным произвести и качественный анализ структуры поведения животного в ходе решения поставленной перед ним задачи. Дрессировка Если при оперантном обусловливании животному предоставляется максимальная возможность проявить инициативу, самостоятельно выбрать способ действия в ходе решения задач, то при дрессировке, наоборот, навыки вырабатываются всецело под целенаправленным воздействием человека и в соответствии с его замыслом. Дрессировка осуществляется путем систематической тренировки животного, при которой подкрепляются требуемые двигательные реакции и их сочетания и упраздняются нежелательные движения. В результате дрессировки, как правило, вырабатываются прочные и четко скоординированные двигательные акты, достигающие подчас большой сложности. Выполняются они животным всегда в ответ на подаваемые человеком сигналы. При цирковой дрессировке, рассчитанной на зрелищный эффект, раньше практиковался преимущественно болевой метод — наказание (иногда весьма жестокое) животного за каждое неправильное движение. В настоящее время его в основном сменил гуманный (и более эффективный) метод дрессировки, основанный на учете особенностей естественного поведения животного, его предварительном приручении, хорошем с ним обращении и пищевом подкреплении правильно выполняемых движений. Иногда применяется смешанный метод — поощрение правильных и наказание за неправильные движения. Дрессировка находит также широкое практическое применение во многих сферах хозяйственной и военной деятельности человека (служебное и охотничье собаководство). В научных исследованиях широко пользуются разными формами дрессировки для изучения различных аспектов психической деятельности животных. Четкость условий, в которые ставится животное, и возможность точного учета сигнальных раздражителей (подаваемых экспериментатором сигналов) делают научную дрессировку незаменимой при изучении навыков животных. Дрессировка является значительно более сложным процессом, чем простое обусловливание, и не сводится лишь к выработке цепей условных рефлексов. Специфическая трудность заключается в том, чтобы дать животному понять, что от него требуется, какие движения оно должно выполнить. Между тем эти движения хотя и входят в видотипичный репертуар поведения, являются зачастую непривычными или трудноосуществимыми в заданных дрессировщиком условиях. Советский зоопсихолог МА.Герд, детально разработавшая теорию дрессировки, делит поэтому процесс дрессировки на три стадии: наталкивания, отработки и упрочения. На первой стадии решается задача — вызвать впервые ту систему движений, которая нужна человеку, «натолкнуть» животное на ее выполнение. Это достигается тремя путями: путем непосредственного наталкивания, косвенного наталкивания или сложного. В первом случае дрессировщик заставляет животное следовать или поворачиваться вслед за пищевым или иным привлекательным для животного объектом. Во втором случае провоцируются движения, непосредственно не направленные на приманку, но обусловливаемые общим возбуждением животного (так формируются преимущественно манипуляционные действия, выполняемые конечностями: обхватывание, перенос, удары или царапанье лапами и т. д.). Так, например, для создания циркового номера (раскатывание ковра лисицей) дрессировщик, стоя около свернутого ковра, поддразнивает ее куском мяса, но не дает схватить его. Возбужденное животное подскакивает, привстает, перебирает передними лапами и т. д. Все случайные прикосновения к ковру при этом закрепляются небольшими кусками пищи, в результате чего лисица все чаще будет обращаться к ковру, и наконец, появятся нужные дрессировщику движения лапой по ковру. В дальнейшем эти движения отрабатываются, направляются на середину рулона и т. д. При сложном наталкивании, по Герд, дрессировщик вначале вырабатывает у животного определенный навык, а затем изменяет ситуацию, заставляя животное по-новому применять выработанное умение. Так, балансирование мячом на кончике носа вырабатывается у морских львов после того, как они научились сбрасывать его в руки дрессировщику. Убирая руки из поля зрения зверя, пряча их за спину, дрессировщик заставляет его несколько задерживать мяч на кончике носа, ибо подкрепление (рыба) будет получено животным лишь после того, как мяч окажется в руках человека. Путем обильного подкрепления постепенно увеличивается длительность удерживания мяча, и в конечном итоге получается знаменитый коронный цирковой номер. Вторую стадию дрессировки, стадию отработки, Герд определяет как этап, на котором совершается отсечение многих лишних движений, вначале сопровождающих необходимые действия животного; далее — отшлифовка первичной, еще весьма несовершенной системы движения и, наконец, выработка удобной сигнализации, с помощью которой дрессировщик в дальнейшем управляет поведением животного. Усилия дрессировщика на этой стадии направлены на упразднение ориентировочных реакций, движений, обусловленных страхом, и иных помех, а также на упорядочивание последовательности, направленности и длительности вырабатываемых движений. Необходимо также заменить реакцию на пищу реакцией на подаваемый дрессировщиком сигнал. При всем этом вновь используются некоторые приемы наталкивания. Например, чтобы отшлифовать у медведя удерживание бутафорного «торта», используется «дробное» наталкивание: дрессировщик поднимает пищевую приманку на нужную высоту и относит ее чуть вбок, в результате чего коробка, которая раньше прижималась медведем к низу живота, поднимается им на уровень груди и немного в сторону. Это правильное положение фиксируется минимальным или средним подкреплением. Равным образом правильная осанка медведя фиксируется приманкой, удерживаемой над его головой, и т. д. С помощью наталкивающих воздействий; производится и выработка искусственной сигнализации. Заключительная стадия процесса дрессировки, стадия упрочения, характеризуется усилиями дрессировщика, направленными на закрепление выработанного навыка и надежность его воспроизведения в ответ на подаваемые сигналы. Дробное наталкивание (приманивание) применяется уже крайне редко, а пищевое подкрепление осуществляется; уже не после нужного элемента, навыка, а преимущественно после целого комплекса выполненных движений. Вообще кормление производится реже, на более крупными порциями. Выработанные в результате навыки приобретают стереотипную форму при которой конец одного действия может послужить началу последующего. Произведенный Герд анализ процесса дрессировки убедительно показывает исключительную сложность, гетерогенность в многоплановость поведения животных при выработке у них искусственный навыков. Но не менее сложная картина обнаруживается и для формировании навыков у животных и в естественных условиях. Об этом всегда следует помнить при ознакомлении с лабораторными экспериментами, где предельное искусственное упрощение и схематизация форм поведения животных являются залогом успеха научного исследования. Познавательные процессы при формировании навыков Еще в начале нашего века сложилось мнение, что образование навыков — как в отношении ориентации среди элементов среды, так в отношении формирования новых сочетаний движений — происходит путем «проб и ошибок». К этому выводу пришел в результате своих исследований ряд выдающихся ученых — Г. Спенсер, К. Ллойд-Морган, Г. Дженнингс, и, прежде всего Э. Торндайк. Согласно концепции «проб и ошибок», животное запоминает то, что случайно привело к успеху, все остальное постепенно отсеивается. Иными словами, в результате «проб и ошибок» совершается отбор и закрепление случайно произведенных удачных движений, что и приводит в конце концов путем многократных повторений к формированию двигательного навыка. Конечно, при этом отсутствует какое бы то ни было понимание связей и отношений между компонентами научения. Существенным здесь является представление, что «пробы и ошибки» совершаются беспорядочно. Однако, как уже было доказано, образование навыков является значительно более сложным процессом и определяется активным отношением животного к воздействующим на него факторам среды. Еще в 20-е годы Э. Толмен, В. П. Протопопов и другие возражали против представления о хаотичности движений, производимых животными при решении задач, и показали, что эти движения формируются в процессе активной ориентировочной деятельности. Пои этом животное анализирует ситуацию и избирает то направление движений, которое соответствует положению «цели». В результате движения животного становятся все более адекватными ситуации, в которой дана задача. Таким образом, на место случайного возникновения движений ставится, как решающий фактор, активный двигательный анализ ситуации. Этот взгляд получил подтверждение в ряде экспериментальных исследований. Так, американский ученый И. Ф. Дэшиелл показал, что пробные забеги крыс в тупики лабиринта вовсе не случайны, а, как правило, производятся в сторону «цели»: после первой ориентировки в лабиринте животное создает как бы общую систему направления своего передвижения; при этом крыса значительно чаще заходит в тупики, расположенные по направлению к цели, чем расположенные в обратном направлении (рис. 12). Аналогичные данные были получены К. Спенсом и В. Шипли (рис. 13). Направленность действия при выработке навыков, возникающая в результате первоначальной активной ориентировки животного, побудила И. Кречевского выдвинуть тезис о появлении у животного своего рода «гипотез», которыми оно руководствуется при решении задач. Особенно это выявляется в тех случаях, если подопытному животному ставится заведомо неразрешимая задача, например, когда в беспорядочной последовательности закрываются и открываются проходы лабиринта (рис. 14). В этом случае у разных групп подопытных животных появляются разные, но всегда устойчивые типы поведения. По Кречевскому, животные пытаются выйти из затруднения, строя «гипотезу» и испытывая ее пригодность. В случае неудачи животное заменяет ее другой «гипотезой». Поэтому действия по одной «гипотезе» повторяются многократно, до выявления ее непригодности. Соответственна животное некоторое время ведет себя одинаковым образом независимо от меняющихся внешних условий. Так, в упомянутом лабиринте Кречевского которые крысы, например, первоначально сворачивали на всех развилках в одну и ту же сторону. Убедившись, что эта «гипотеза» не приводит к успеху, они стали постоянно сворачивать в противоположную сторону.  Рис. 12. Лабиринт Дэшиелла. Стрелка показывает вход в лабиринт, пунктирные линии обозначают дверцы, открывающиеся только в сторону выхода из лабиринта. Из 6 Т-образных тупиков наиболее часто посещался крысами тупик В, расположенный по направлению к цели (параллельно выходу). Тупики Б,Г и Е, направленные в противоположную от цели сторону, посещались в 5–11 раз реже, чем тупик В Рис. 12. Лабиринт Дэшиелла. Стрелка показывает вход в лабиринт, пунктирные линии обозначают дверцы, открывающиеся только в сторону выхода из лабиринта. Из 6 Т-образных тупиков наиболее часто посещался крысами тупик В, расположенный по направлению к цели (параллельно выходу). Тупики Б,Г и Е, направленные в противоположную от цели сторону, посещались в 5–11 раз реже, чем тупик В  Рис. 13. Лабиринт Спенса и Шипли. П — пусковая камера; В — выход из лабиринта. Пунктирные линии обозначают непрозрачные занавесы, не дающие крысе увидеть концы коридоров. Тупики, расположенные в сторону выхода, посещались в 3–4 раза чаще, чем направленные в противоположную сторону Рис. 13. Лабиринт Спенса и Шипли. П — пусковая камера; В — выход из лабиринта. Пунктирные линии обозначают непрозрачные занавесы, не дающие крысе увидеть концы коридоров. Тупики, расположенные в сторону выхода, посещались в 3–4 раза чаще, чем направленные в противоположную сторону В других случаях крысы начинали регулярно чередовать повороты налево и направо. Таким образом, обнаруживается четкая связь между предыдущей попыткой и последующей, животное как бы стремится организовать свое поведение по одному «принципу». Кречевский считал, что этот в известней мере абстрактный «принцип» отличается систематичностью и обусловлен внутренней «настройкой» животного.  Рис. 14. Лабиринт Кречевского. П — пусковая камера. Прерывистыми линиями показаны дверцы, которыми можно закрывать проход то с левой, то с правой стороны, пунктиром — путь животного. Пояснения см. в тексте Рис. 14. Лабиринт Кречевского. П — пусковая камера. Прерывистыми линиями показаны дверцы, которыми можно закрывать проход то с левой, то с правой стороны, пунктиром — путь животного. Пояснения см. в тексте Концепция Кречевского, безусловно, не свободна от недостатков, и сам термин «гипотеза» является крайне неудачным применительно к поведению животных. Вместе с тем большая заслуга этого ученого заключается в том, что он убедительно показал всю сложность поведения крысы в лабиринте (и других подобных ситуациях), особенно в начальном периоде решения задачи, когда решающую роль играет исследовательское поведение животного. Не менее ценным является в этой концепции то, что делается акцент на активность и инициативу животного и подчеркивается роль внутренних факторов (психического настроя животного) в процессе решения задач. Опровергается тезис о «случайности» попыток решения задач и опытами с применением «латентного обучения». В таких опытах сравнивается скорость образования навыка у животных, которых непосредственно перед началом эксперимента помещают в лабиринт или «проблемный» ящик, с таковой у животных, которым предоставлялась возможность предварительно активно ознакомиться с установкой, побегать в ней. В последнем случае вырабатывается двигательный навык значительно быстрее. В этой связи важно отметить, что каждая крыса, впервые попавшая в лабиринт, ведет себя иначе, особенно на первоначальной ориентировочной фазе, предшествующей решению задачи, когда пробежки совершаются еще без всякого подкрепления и служат лишь накоплению опыта. Варьирует даже модальность ведущей рецепции при первоначальном обследовании лабиринта (одни крысы руководствуются преимущественно визуальными, другие — кинестетическими стимулами и т. д.). Крыса, просто бегая по лабиринту, познает его еще до решения задачи, связанного с получением пищевого подкрепления (или наказания) за правильную (или неправильную) ориентацию. Разумеется, индивидуальные особенности исследовательского поведения животного сказываются и в ходе самого решения задачи. Значительные индивидуальные различия поведения вообще являются характерным признаком процессов научения. Итак, активная познавательная деятельность животного является важнейшей предпосылкой успешного формирования навыка при решении задач. В сущности, именно познавательный компонент определяет природу навыка. По Леонтьеву, важнейшим критерием навыка является выделение при решении задач особого состава или стороны деятельности, отвечающей условиям, в которых дан побуждающий деятельность животного предмет. Леонтьев обозначил этот компонент деятельности операцией. Выделение операции в двигательной активности животного и указывает на то, что мы действительно имеем дело с истинным навыком, а не с другой формой научения, поэтому Леонтьев считает навыками лишь закрепленные операции. Выделение операции можно показать на простом опыте, проведенным А. В. Запорожцем и И. Г. Диманштейн по методу «обходного пути». В аквариум ставится поперечная перегородка из марли так, чтобы у боковой стенки оставался свободный проход. В меньшую часть аквариума в начале опыта помещается подопытная рыба, в большую, за перегородкой, — приманка (кусочки мяса). Чтобы заполучить приманку, подопытное животное должно обойти преграду (перегородку), что ему и удается после ряда безуспешных попыток найти дорогу к пище напрямик. В поисках пути к приманке животное производит локомоторные действия, в которых Леонтьев вычленяет двоякое содержание: 1) направленную деятельность, приводящую к результату и возникающую под влиянием свойства самого побуждающего деятельность предмета (запахи мяса), и 2) деятельность, связанную с воздействием преграды, т. е. с условиями, в которых дан побуждающий деятельность предмет. Именно эта деятельность и явится операцией. После того как рыба (в описанном опыте — сомик) хорошо усвоила обходной путь, преграда была удалена, тем не менее рыба полностью повторяла прежний путь, как если бы преграда была на месте. Только постепенно путь рыбы к приманке выпрямлялся (рис. 15,а). Подобные опыты производились со сходными результатами и на крысах (рис. 15, 6). Следовательно, отмеченные компоненты деятельности здесь выступают еще слитно; воздействие, определяющее обходное движение, еще прочно связывается с воздействием пищи, с ее запахом. Перегородка не отделяется от приманки, воздействие преграды пока еще не воспринимается как свойство другой вещи — словом, операция здесь еще не выделяется. По поводу описанного здесь опыта с сомиком Леонтьев пишет, что деятельность животных определяется уже фактически воздействием со стороны отдельных вещей (пища, преграда), но отражение действительности остается у них отражением совокупности отдельных ее свойств. В приведенном примере познавательный компонент, а соответственно и навык в целом находятся на еще очень низком уровне; выученная животным траектория обходного пути оказалась настолько прочно закрепленной, что лишь постепенно элиминировалась в новых условиях среды (т. е. после удаления перегородки). Это типичный пример автоматизированного навыка. Чем сложнее навык, тем больше его познавательное значение и наоборот. В навыках высшего ранга, характерных для высших позвоночных, операция играет чрезвычайно важную познавательную роль. Однако и у этих животных закрепление индивидуального опыта совершается нередко в форме примитивных навыков наподобие описанного. Уровень навыка зависит в каждом конкретном случае от определяющих факторов биологии вида и от ситуации, в которой животное встречается с той или иной более или менее сложной задачей. 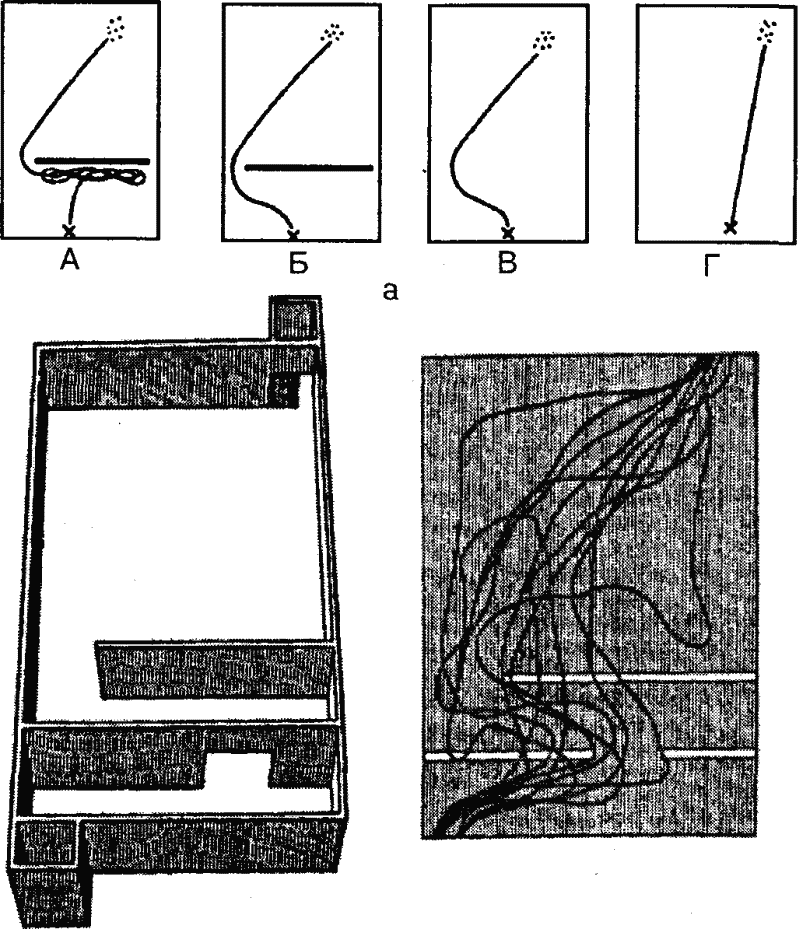 Рис. 15. Формирование обходного пути и его сохранение после удаления преград у сомика (а) и крысы (б). Пояснения см. в тексте Рис. 15. Формирование обходного пути и его сохранение после удаления преград у сомика (а) и крысы (б). Пояснения см. в тексте Говоря о структуре и динамике навыка, важно подчеркнуть, что понятие преграды не следует понимать лишь в физическом смысле, как перегородку на пути животного. Под преградой понимается любое препятствие к достижению побуждающего объекта («цели») при решении задачи. Это подчеркивал в свое время и Протопопов, который сумел экспериментально доказать, что любые двигательные навыки формируются у животных путем преодоления «преграды», что содержание навыков определяется именно характером самой преграды. Стимул же влияет, по Протопопову, на навык только динамически, определяя быстроту и прочность его закрепления. Таким образом, преодоление преграды составляет наиболее существенный элемент формирования навыка не только при его. выработке описанным методом, но и при всех других методах, широко применяемых в зоопсихологических исследованиях. В частности, это относится к методам «лабиринта» и «проблемного ящика». Именно в способах преодоления преграды проявляется познавательная функция навыка. Изучая познавательные аспекты формирования навыков у животных, венгерский зоопсихолог Л. Кардош также показал, что в ходе обучения в лабиринте у животного накапливается значительный запас информации. В результате уже в начале лабиринта животное «в памяти… видит дальше стен, закрывающих его поле ощущений; эти стены становятся как бы прозрачными. В памяти оно „видит“ цель и наиболее важные с точки зрения локомоции (перемещения) части пути, открытые и закрытые двери, разветвления и прочее, „видит“ точно так и там, где и как оно видело и в действительности во время обхода лабиринта».[36] 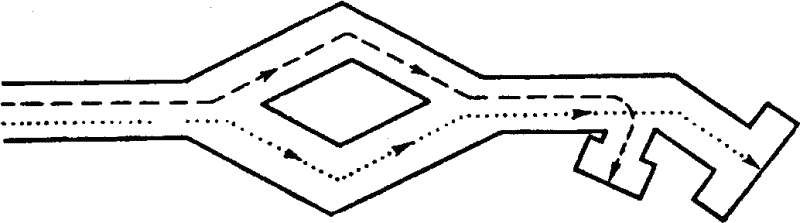 Рис. 16. Схема опыта Кардоша. Пояснения см. в тексте Рис. 16. Схема опыта Кардоша. Пояснения см. в тексте Вместе с тем Кардош четко показал и границы познавательных возможностей животного в лабиринте, как и вообще при решении пространственно временных задач. Здесь имеются две возможности: локомоторное и манипуляционное познавание (второе происходит при формировании инструментальных навыков). В первом случае животное изменяет свое пространственное отношение к среде без того, чтобы сама среда изменилась. Если же в окружающей животное среде что-то изменяется в результате изменения поведения животного, то речь идет уже о манипуляторной активности животного. В исследованиях, произведенных вместе с И. Баркоци, Кардош показал, что с большим трудом можно крысу научить выбирать в одном и том же лабиринте разные пути, приводящие к одной точке, а затем двигаться дальше различным образом, например прямо или в сторону (рис. 16). Это пример локомоторного познавания. Но, согласно Кардошу, нельзя обучить животное (за исключением, возможно, человекообразных обезьян) тому, что в зависимости от выбора того или иного пути передвижения «случается то или нечто другое», т. е. произойдут изменения в окружающей среде (в экспериментах пища подменялась другим подкреплением — водой). Человек же, как пишет Кардош, «удивился бы, найдя в одном и том же месте разные предметы, когда он подходил справа и слева, но он обучился бы после первого же опыта. Развитие именно здесь делает скачок… Человек полностью может освободиться от направляющего воздействия пространственного порядка, если временно-причинные связи требуют другого».[37] Научение и общение. Подражание Существенную роль в формировании поведения высших животных играют явления подражания, которые в основном, хотя и не все, относятся к сфере научения. Мы здесь не будем рассматривать формы подражания, которые относятся к инстинктивному поведению. Это широко распространенная среди животных взаимная стимуляция — аллеломиметическое поведение (рис. 17), при котором выполнение видотипичных действий одними животными является побуждающим фактором для других. Последние в результате начинают выполнять такие же действия (одновременные отдых, собирание пищи и т. д.). Следовательно, в данном случае имеет место взаимопоощрение видотипичной деятельности. Научение же путем подражания («имитационное научение») заключается в индивидуальном формировании новых форм поведения, но путем одного лишь непосредственного восприятия действий других животных. Таким образом, мы имеем здесь дело с научением на основе общения. Имитационное научение, как и всякое научение вообще, можно подразделить на облигатное и факультативное. При облигатном имитационном научении результат научения вполне укладывается в рамки видового стереотипа. Особенно это относится к молодым животным, которые путем подражания научаются выполнять некоторые жизненно необходимые действия обычного поведенческого «репертуара» своего вида. Так, у молоди стайных рыб защитная реакция на появление хищника (бегство) формируется в результате подражания поведению других рыб при одном лишь виде поедания хищником членов стаи. Л. А. Орбели считал такое имитационное поведение «главным охранителем вида», ибо «громадное преимущество заключается в том, что „зрители“, присутствующие при акте повреждения члена их же стада или их сообщества, вырабатывают рефлекторные защитные акты и таким образом могут в будущем избежать опасности».[38] Облигатное имитационное научение является также важным элементом реакции следования (см. ч. II, гл. 3) и распознавания молодыми млекопитающими пищевых объектов. Путем облигатного имитационного научения молодые животные накапливают опыт в гнездостроении у птиц (В. П. Промптов) и шимпанзе (Дж. ван Лавик-Гудолл) и т. д. 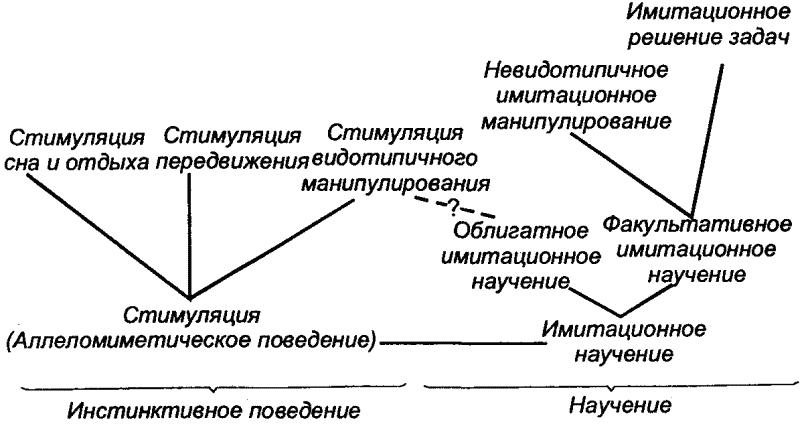 Рис. 17. Схема категории подражания у животных (по Фабри) Рис. 17. Схема категории подражания у животных (по Фабри) Факультативное имитационное научение в простейших формах представлено в имитации невидотипичных движений на основе облигатного (аллеломиметического) стимулирования. Сюда относятся, например, случаи имитирования обезьянами действий человека, особенно при их содержании в домашней обстановке. Производимые ими при этом действия с предметами быта или инструментами, конечно, выходят за рамки видового поведения. Поскольку здесь имеет место научение новым приемам манипулирования, в данном случае можно говорить о невидотипичном имитационном манипулировании. Высшим проявлением факультативного имитационного научения следует явно считать решение задач путем подражания (или хотя бы облегчение решения). При таком «имитационном решении задач» у «животного-зрителя» вырабатывается определенный навык в результате одного лишь созерцания действий другой особи, направленных на решение соответствующей задачи. Способность к этому установлена у разных млекопитающих: человекообразных и низших обезьян, собак, кошек, крыс. У обезьян имитационное решение задач играет, очевидно, особенно большую роль. Советский исследователь поведения животных А. Д. Слоним, например, считает, что формирование условных рефлексов происходит в обезьяньем стаде преимущественно на основе подражания. В пользу этого говорят и полевые наблюдения, выполненные рядом исследователей в последние годы. Правда, на основе имитационного научения у «зрителей», очевидно, не могут формироваться инструментальные навыки. Об этом свидетельствуют, в частности, опыты американского исследователя Б. Б. Бека, в которых павианы-«зрители» наблюдали за употреблением орудия при решении задачи у сородича. «Зрители» не оказались способными к имитационному решению столь сложной задачи, но впоследствии они намного чаще и интенсивнее, чем до опытов, манипулировали этим орудием. Это показывает активирующую роль аллеломиметического поведения и невидотипичного имитационного манипулирования при выработке сложных навыков в условиях общения. Соотношения отдельных категорий и форм подражания у животных можно проиллюстрировать схемой (по К. Э. Фабри — см. рис. 17). Эта схема неполная в том отношении, что не отражает явления подражания в области сигнализации и коммуникации (например, звукоподражание у птиц). Однако здесь нет принципиальных отличий от отмеченных общих закономерностей и характеристик, и поэтому данная классификация в соответствующих своих разделах вполне применима и для этой области. Так, например, к стимуляции видотипичного поведения — относится и стимуляция видотипичной акустической сигнализации («хоровое пение» у лягушек или птиц), невидотипичному имитационному манипулированию соответствует подражание чужим звукам и песням у птиц, а к облигатному имитационному научению относится, в частности, освоение птенцами видотипичных звуков путем подражания пению взрослых особей. Еще в 1773 г. английский ученый Д. Баррингтон воспитывал молодых коноплянок совместно с птенцами жаворонков. В результате коноплянки перенимали пение жаворонков. С тех пор аналогичные опыты ставились на птицах многократно. Что же касается обезьян, то в опытах, поставленных Н. А. Тих на молодых гамадрилах, намечались, правда нечетко, некоторые намеки на возможность выработки даже таких навыков, которые можно было бы квалифицировать как имитационное решение задач на основе звукоподражания (в этих опытах обезьяны получали пищевое подкрепление за подражание звукам, которым в их присутствии обучали других обезьян). Исходя из приведенной классификации можно определить два принципиально различных подхода к изучению подражания у животных. Аллеломиметическое поведение изучается путем раздельного обучения изолированных друг от друга животных, которые затем сводятся вместе. Советский зоопсихолог Б. И. Хотин, например, обучал одних животных (рыб, птиц, млекопитающих) реагировать положительно на определенный сигнал, а других — отрицательно. После того как животные были сведены вместе и подвергнуты воздействию этого раздражителя, выяснилось, что преобладает: результаты обычного (неимитационного) научения или взаимная стимуляция. В результате можно судить о силе аллеломиметической реакции, т. е. инстинктивного подражания. При изучении же имитационного научения животные с самого начала общаются между собой. Одна особь (или несколько особей) выполняет при этом роль «актера», т. е. обучается экспериментатором за соответствующее вознаграждение на виду у других особей — «зрителей». Если в результате последние без собственных упражнений и подкрепления вследствие одних лишь наблюдений за действием «актера» также научаются решать поставленную ему задачу, то можно говорить о факультативном имитационном научении и даже об имитационном решении задач. Так, например, в экспериментах с обезьянами все участвующие в опыте животные устремляются к месту его проведения и внимательно наблюдают, как вожак добывает и получает подкрепление. В итоге соответствующие навыки формируются у всех «зрителей», что проявляется в успешном решении ими той же задачи в отсутствие вожака. На этом примере видно, как проявляется еще одна особенность, характерная для проявлений подражания в естественных условиях. Дело в том, что наряду с взаимным стимулированием к совместному выполнению определенных (инстинктивных) действий в сообществах животных действует и фактор противоположного рода — подавление действий членов сообщества «господствующими» особями. Так, в приведенном примере обезьяны боялись вплотную подойти к экспериментальной установке, а тем более брать пищу из кормушки. Вместе с тем у обезьян существуют особые «примиряющие» сигналы, оповещающие доминирующую особь о готовности «зрителей» «только смотреть», чем обеспечивается возможность осуществления аллеломиметического поведения и имитационного научения. Отсюда явствует, что явления подражания сложным образом переплетаются с внутригрупповыми отношениями животных. * * *Подводя итог обзору инстинктивного поведения и научения у животных, вернемся еще раз к описанной ранее общей схеме поведенческого акта. Эта схема, разумеется, не отражает всего многообразия вариантов поведенческих актов в их конкретных проявлениях. Но во всех этих вариантах остается в силе основная закономерность: каждое действие животного начинается с внутреннего стимула, который в виде потребности активирует животное, кладет начало его поиску раздражителей. Это начало, как и общее направление этого поиска, всегда генетически фиксировано, равно как и конец — заключительные движения животного в завершающей фазе как пройти этот путь от начала до конца между этими «жестко запрограммированными» основными элементами поведения — зависит от лабильных компонентов поведения животного, от его умения быстро и правильно ориентироваться во всех конкретных ситуациях, в которые оно попадает. Как мы видели, особое значение это имеет в начале пути. У высших животных факультативное научение, сопряженное с формированием операций, является основным средством для продвижения к завершающей фазе. Но поскольку операция направлена на преодоление преграды, т. е. условий, в которых дан объект завершающего поведения, то здесь особенно важна более полноценная по возможности ориентировка животного во времени и пространстве. Ясно, что эта ориентировка будет тем более совершенной, чем выше уровень психической деятельности животного, его познавательные способности. Наибольший эффект обеспечивают в этом отношении высшие психические функции, особенно интеллектуальные способности, придающие всему поведению животного наибольшую гибкость и адаптивность. К этому вопросу мы еще вернемся при рассмотрении эволюции психики. Примечания:1 2-е издание учебника подготовила полностью к публикации ученица К. Э. Фабри — кандидат психологических наук Н. Н. Мешкова. 2 Герцен А. И. Избранные философские сочинения. М., 1940. С. 222–223. 3 Выготский Л. С. Развитие высших психических функций. М., 1960. С. 405. 12 Ламарк. Философия зоологии. Т. 2. М.; Л., 1937. С. 325. 13 Там же. 14 Там же. 16 Там же. С. 326. 17 Там же. С. 327. 18 Рулье К. Ф. Избранные биологические произведения. М., 1954. С. 317. 19 Там же. С. 332. 20 Там же. 21 Там же. С. 317. 22 Там же. С. 441. 23 Дарвин Ч. Происхождение видов. М., 1952. С. 260. 24 Там же. 25 Там же. С.261. 26 Дарвин Ч. Избранные письма. М., 1950. С. 106. 27 Дарвин Ч. Происхождение видов. С. 261. 28 Там же. С.278. 29 Там же. С.284. 30 Там же. С. 283. 31 Там же. 32 Дарвин Ч. Происхождение человека и половой отбор. Спб., 1896. С. 45. 33 Там же. 34 Северцов А. Н. Собр. соч. Т. III. M.; Л., 1945. С. 300. 35 Скребицкий Г. Отношение чайки к гнезду, яйцам и птенцам // Рефлексы, инстинкты и навыки. М., 1936. С. 144–145. 36 Кардош Л. Некоторые проблемы сравнительно-психологического анализа процесса учения // Annales Universitatis Scientiarum Budapestinensis. Sect. Paedagogica et Psychologica. T.1. Budapest, 1970. P. 15. 37 Ibid. P. 19. 38 Орбели Л. А. Вопросы высшей нервной деятельности. М.; Л., 1949. С. 352. |
|
||