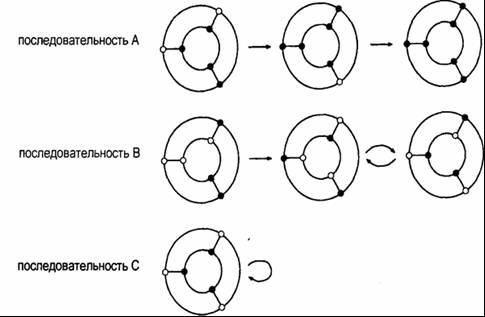
|
||||
|
|
Часть IV Природа жизни Глава 7 Новый синтез 'Теперь мы можем вернуться к центральному вопросу этой книги: что есть Жизнь? Мой тезис заключался в том, что в настоящее время зарождается теория живых систем, совместимая с философскими основами глубокой экологии, включая соответствующий математический язык и немеханистическое посткартезианское понимание Жизни. Паттерн и структура Возникновение и уточнение понятия паттерн организации было исключительно важным этапом в развитии нового способа мышления. От Пифагора и Аристотеля до Гете и организменных биологов лежит непрерывная интеллектуальная традиция: ученые стремятся понять паттерн, сознавая, что это чрезвычайно важно для понимания живой формы. Александр Богданов первым попытался объединить понятия организации, паттерна и сложности в последовательную теорию систем. Кибернетики сосредоточились на паттернах связи и управления — в частности, на паттернах круговой причинности, лежащих в основе концепции обратной связи; благодаря этому, они первыми четко разграничили паттерн организации системы и ее физическую структуру. За последние двадцать лет были найдены и проанализированы недостающие «элементы головоломки» — концепция самоорганизации и новая математика сложных систем. И снова понятие паттерна оказалось центральным в обоих этих направлениях. Концепция самоорганизации возникла из осознания сети как общего паттерна жизни; эта концепция в дальнейшем была развита Матураной и Варелой в их теории автопоэза. Новая математика сложных систем представляет собой, по существу, математику визуальных паттернов — странных аттракторов, фазовых портретов, фракталов и т. п., — которые анализируются в контексте топологической структуры, впервые разработанной Пуанкаре. Понимание паттерна, таким образом, приобретает решающее значение в научной концепции жизни. Тем не менее при всей своей важности, для полного понимания живой системы оно недостаточно. Мы должны понять также структуру системы. Мы уже знаем, что изучение структуры было основной целью западной науки и философии и как таковое снова и снова отодвигало на второй план изучение паттерна. Я пришел к убеждению, что ключ к построению полной теории живых систем заложен в синтезе двух подходов, т. е. в едином изучении паттерна (или формы, порядка, качества) и структуры (или вещества, материи, количества). Я буду следовать за Умберто Матураной и Франциско Варелой в их определении этих двух ключевых критериев живой системы — ее паттерна организации и ее структуры'. Паттерн организации любой системы, живой или неживой, — это конфигурация взаимоотношений между компонентами системы, определяющая существенные характеристики этой системы. Другими словами, необходимо наличие определенных взаимоотношений, чтобы данный объект мог быть признан, скажем, стулом, велосипедом или деревом. Именно ту конфигурацию взаимоотношений, которая обусловливает существенные характеристики системы, мы и будем понимать как паттерн организации. Структура системы — это физическое воплощение ее паттерна организации. Если описание паттерна организации означает абстрактное отображение взаимоотношений, то описание структуры включает характеристики реальных физических компонентов системы — их форму, химический состав и т. п. Чтобы проиллюстрировать разницу между паттерном и структурой, рассмотрим хорошо знакомую неодушевленную систему — велосипед. Для того чтобы нечто можно было назвать велосипедом, в нем должны существовать определенные функциональные взаимоотношения между компонентами, известными как рама, педали, руль, колеса, цепное колесо и т. п. Полная конфигурация этих функциональных взаимоотношений и составляет паттерн организации велосипеда. Необходимо наличие всех этих взаимоотношений, чтобы система обладала существенными характеристиками велосипеда. Структура велосипеда представляет собой физическое воплощение его паттерна организации в виде компонентов конкретной формы, изготовленных из конкретных материалов. Один и тот же паттерн велосипеда может быть воплощен во множестве различных структур велосипеда. Рули имеют различную форму для прогулочного велосипеда, гоночного велосипеда или горного велосипеда; рама может быть тяжелой и твердой или легкой и тонкой; покрышки могут быть узкими или широкими, надувными или изготовленными из сплошной резины. Все эти комбинации и множество других легко распознаются как различные структурные воплощения одного и того же паттерна взаимоотношений, определяющего велосипед. Три ключевых критерия Для машины, подобной велосипеду, отдельные ее части проектируются, изготавливаются и затем собираются воедино, образуя структуру с фиксированными компонентами. В отличие от такой структуры, в живой системе компоненты непрерывно меняются. Через живой организм идет непрерывный поток материи. Каждая клетка постоянно синтезирует и растворяет структуры, а также удаляет отработанные продукты. Клетки тканей и органов заменяются в процессе непрерывных циклов. Идет рост, развитие и эволюция. Таким образом, с самого зарождения биологии понимание живой структуры было неотделимо от понимания метаболических и эволюционных процессов2. Это поразительное свойство живых систем предполагает процесс как третий критерий полного описания природы жизни. Жизненный процесс — это деятельность, направленная на постоянное воплощение и поддержание паттерна организации системы. Таким образом, процесс служит связующим звеном между паттерном и структурой. В случае велосипеда, паттерн организации представлен чертежами конструкции, которые используются при изготовлении велосипеда, структура — это конкретный материальный велосипед, а связующее звено между паттерном и структурой находится в мозгу конструктора. В живом же организме паттерн организации всегда воплощен в структуре организма, а связующим звеном между паттерном и структурой служит процесс постоянного воплощения. Критерий процесса завершает концептуальную структуру моего синтеза зарождающейся теории живых систем. Определения этих трех критериев — паттерна, структуры и процесса — еще раз приводятся в таблице, представленной ниже. Все три критерия полностью взаимозависимы. Паттерн организации может быть распознан только при том условии, что он воплощен в физическую структуру — а в живых системах это воплощение является непрерывным процессом. Таким образом, структура и процесс неразрывно связаны. Можно сказать, что три критерия — паттерн, структура и процесс — это три различные, но неразделимые точки зрения на феномен жизни. Они образуют три концептуальных измерения моего синтеза. Понять природу жизни с системной точки зрения означает определить набор общих критериев, по которым можно провести четкое различие между живыми и неживыми системами. За всю историю биологии было предложено много критериев, но все они по той или иной причине оказывались неадекватными. Тем не менее последние формулировки модели самоорганизации и математика сложных систем показывают, что сегодня определить такие критерии возможно. Ключевая идея моего синтеза состоит в том, чтобы выразить эти критерии в рамках трех концептуальных измерений — паттерна, структуры и процесса. Ключевые критерии живой системы паттерн организации конфигурация взаимоотношений, определяющая существенные характеристики системы структура физическое воплощение паттерна организации системы жизненный процесс деятельность, направленная на непрерывное воплощение паттерна организации системы По сути, я предлагаю понимать автопоэз так, как Матурана и Варела определяют паттерн жизни (т. е. паттерн организации живых систем)3; диссипативную структуру — как Пригожий определяет структуру живых систем4; и обучение — как Грегори Бэйтсон и, более полно, Матурана и Варела определяют жизненный процесс. Паттерн организации определяет существенные характеристики системы. В частности, он определяет, является ли система живой или нет. Автопоэз — паттерн организации живых систем — является, таким образом, определяющей характеристикой жизни в новой теории. Чтобы выяснить, относится ли данная сущность — кристалл, вирус, клетка или Земля — к живым системам, нужно определить одно: является ли ее паттерн организации автопоэзной сетью. Если да, то мы имеем дело с живой системой; если нет, то это — неживая система. Обучение [cognition] (или процесс жизни), как мы увидим ниже, неразрывно связано с автопоэзом. Автопоэз и обучение — это два разных аспекта одного феномена жизни. По новой теории, все живые системы — это когнитивные, т. е. обучающиеся системы, а обучение всегда предполагает существование автопоэзной сети. Что касается второго критерия жизни, структуры живых систем, то здесь ситуация несколько иная. Хотя структура живой системы всегда диссипативна, не все диссипативные структуры являются автопоэзными сетями. То есть диссипативная структура может быть либо живой, либо неживой системой. Например, клетки Бенара и химические часы, подробно исследованные Пригожиным, являются диссипативными структурами, но не живыми системами5. Три ключевых критерия жизни, вместе с теориями, лежащими в их основе, будут подробно рассмотрены в последующих главах. Здесь я даю лишь предварительный краткий обзор. Автопоэз — паттерн жизни Уже в первой четверти столетия было известно, что паттерн организации живой системы всегда является сетевым паттерном6. Однако мы знаем также, что не всякая сеть представляет живую систему. Согласно Матуране и Вареле, определяющей особенностью живой сети служит то, что она непрерывно производит саму себя. Таким образом, «бытие и творение [живых систем] неразделимы, и в этом заключается специфика их организации»7. Автопоэз, или «самосоздание», — это сетевой паттерн, в котором каждый компонент сети участвует в создании или трансформации других компонентов. Таким образом, сеть непрерывно производит, создает саму себя. Она создается своими компонентами и, в свою очередь, создает эти компоненты. Простейшей из известных нам живых систем является клетка, и Матурана и Варела широко использовали биологию клетки при изучении характеристик автопоэзных сетей. Базовый паттерн автопоэза удобно проиллюстрировать на примере клетки растения. На рис. 7–1 изображена упрощенная картина такой клетки: здесь компонентам даны наглядные условные названия. Соответствующие специальные термины, произведенные из греческого и латинского, читатель найдет в словаре, приведенном ниже.
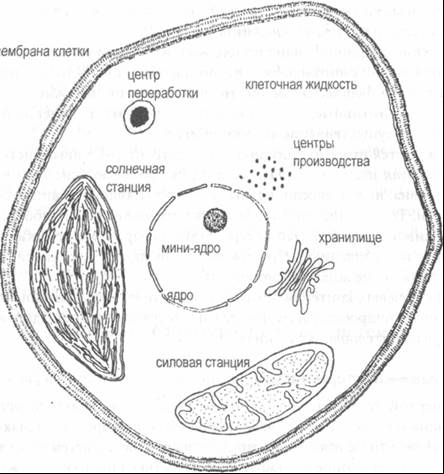
Рис. 7–1. Основные компоненты клетки растения Типичная растительная клетка, как и любая другая, состоит из клеточной мембраны, в которой помещается клеточная жидкость. Жидкость представляет собой густой молекулярный раствор питательных веществ клетки, т. е. химических элементов, из которых клетка строит свои структуры. В клеточной жидкости мы обнаруживаем во взвешенном состоянии ядро клетки, а также большое количество центров производства, где изготавливаются основные структурные строительные блоки, и несколько специализированных частей, называемых «органеллами» — поскольку они аналогичны органам тела. Наиболее важными из этих органелл являются хранилище, центры переработки, силовые и солнечные станции. Как и клетка в целом, ядро и органеллы окружены полупроницаемыми мембранами, которые выборочно пропускают определенные вещества внутрь и наружу. Мембрана клетки, в частности, впускает питательные вещества и рассеивает отходы. Ядро клетки содержит генетический материал — молекулы ДНК, несущие генетическую информацию, и молекулы РНК, которые производятся ДНК и доставляют инструкции в центры производства8. В ядре содержится меньшее «мини-ядро», где создаются производственные центры, которые затем распределяются по всей клетке. Словарь технических терминов клеточная жидкость — цитоплазма («клеточная жидкость») мини-ядро — ядрышко центр производства — рибосома; состоит из рибонуклеиновой кислоты (РНК) и микросомы («микроскопического тела») и представляет собой крошечную гранулу, содержащую ДНК хранилище — аппарат Гольджи (назван по имени итальянского физика Камилло Гольджи) центр переработки — лизосома («растворяющее тело») силовая станция — митохондрия («нитевидная гранула») носитель энергии — аденозинтрифосфат (АТФ), химическое соединение, состоящее из основы, Сахаров и фосфатов солнечная станция — хлоропласт, фотосинтезирующее органическое вещество («зеленый лист») Центры производства представляют собой гранулярные тела, в которых производятся протеины клетки. Последние включают структурные протеины, а также ферменты — катализаторы, содействующие всем молекулярным процессам. В каждой клетке содержится около 500 000 центров производства. Хранилища — это склады плоских мешочков, уложенных примерно как лепешки хлеба-лаваша; здесь хранятся, а затем маркируются, упаковываются и рассылаются по местам назначения различные клеточные продукты. Центры переработки — это органеллы, в которых содержатся ферменты для переваривания пищи, поврежденные компоненты клетки и различные неиспользованные молекулы. Испорченные элементы здесь перерабатываются и используются для построения новых компонентов клетки. Силовые станции выполняют дыхательные функции клетки, т. е. используют кислород для разложения органических молекул на углекислый газ и воду. Отсюда исходит энергия, которая концентрируется в специальных энергетических носителях. Эти энергетические носители представляют собой сложные молекулярные соединения, которые перемещаются к другим частям клетки и снабжают энергией все клеточные процессы, именуемые в совокупности клеточным метаболизмом. Энергетические носители служат основными энергетическими единицами клетки, примерно как деньги в человеческой экономике. Только недавно было обнаружено, что силовые станции содержат собственный генетический материал и делятся независимо от деления клетки. Согласно теории Линн Маргулис, они происходят от простых бактерий, которые поселились в более сложных и крупных клетках примерно два миллиарда лет тому назад9. С тех пор они стали непременными резидентами во всех высших организмах, передаются от поколения к поколению и живут в тесном симбиозе с любой клеткой. Как и силовые станции, солнечные станции имеют собственный генетический материал и самовоспроизводятся, но они содержатся лишь в зеленых растениях. Это центры фотосинтеза, преобразующие солнечную энергию, углекислый газ и воду в сахара и кислород. Произведенные сахара отправляются в силовые станции, где из них извлекается энергия, которая может затем храниться в энергетических носителях. В дополнение к сахарам, растения поглощают также питательные вещества и некоторые другие элементы из земли с помощью корней. Очевидно, что даже для весьма грубого представления о внутриклеточной организации необходимо достаточно сложное описание компонентов клетки; сложность неизмеримо возрастает, когда мы пытаемся представить огромную сеть этих компонентов и их взаимосвязи, означающие тысячи метаболических процессов. Одни только ферменты образуют запутанную сеть каталитических реакций, поддерживающих все метаболические процессы; чтобы обеспечивать их горючим, соответствующую энергетическую сеть составляют энергетические носители. На рис. 7–2 еще раз изображена упрощенная схема клетки растения, но на этот раз стрелками показаны некоторые важнейшие связи в сети метаболических процессов.
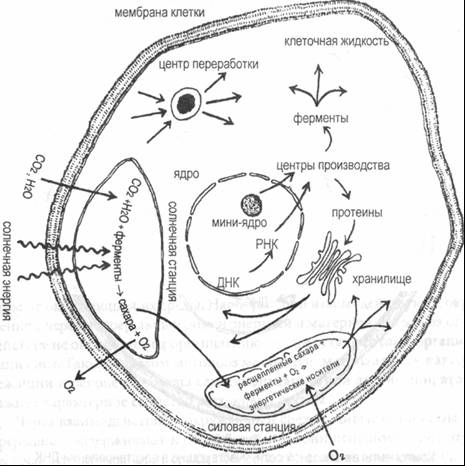
Рис. 7–2. Метаболические процессы в клетке растения Чтобы проиллюстрировать природу этой сети, рассмотрим только одну петлю. ДНК в ядре клетки производит молекулы РНК, которые содержат инструкции для производства протеинов, включая ферменты. Среди последних есть группа специальных ферментов, которые могут распознавать, устранять и заменять поврежденные участки ДНК10. На рис. 7–3 представлена схема некоторых взаимоотношений в такой петле. ДНК производит РНК, которая доставляет инструкции по производству ферментов в центры производства ферментов; произведенные ферменты проникают в ядро клетки и там восстанавливают ДНК. Каждый компонент этой небольшой сети участвует в производстве или преобразовании других компонентов; эта сеть, таким образом, явно обладает признаками автопоэза: ДНК производит РНК; РНК определяет ферменты; а ферменты восстанавливают ДНК.
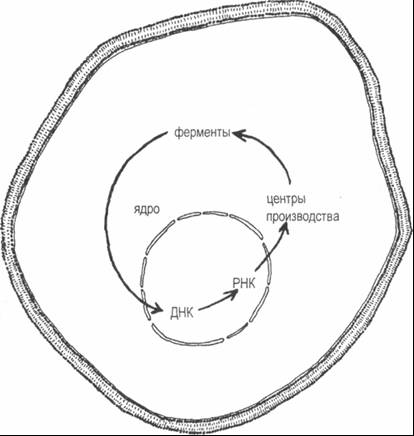
Рис. 7–3. Компоненты автопоэзной сети, участвующие в восстановлении ДНК Чтобы завершить картину, необходимо добавить строительные блоки, из которых построены ДНК, РНК и ферменты; энергетические носители, подающие топливо для всех изображенных процессов; генерацию энергии на силовых станциях на основе расщепленных Сахаров; производство Сахаров в процессе фотосинтеза на солнечных станциях; и т. д. и т. п. С каждым новым добавлением мы убеждаемся, что новые компоненты также помогают производить или трансформировать другие компоненты и что, таким образом, автопоэзная, самосозидающая природа всей сети становится все более очевидной. Особенно интересна клеточная мембрана. Это граница клетки, образованная некоторыми компонентами клетки; она охватывает всю сеть метаболических процессов и тем самым ограничивает их распространение. Вместе с тем мембрана участвует в этой же сети: с помощью специальных фильтров она отбирает сырье для процессов производства (пищу клетки), а отходы производства выводит во внешнюю среду. Таким образом, автопоэзная сеть создает свою собственную границу, которая определяет клетку как отчетливую систему и в то же время сама остается активной частью сети. Поскольку каждый компонент автопоэзной сети производится другими компонентами этой же сети, вся система организационно закрыта; вместе с тем она открыта по отношению к потоку энергии и материи. Организационная закрытость означает, что живая система является самоорганизующейся в том смысле, что ее порядок и поведение не обусловлены окружением, но устанавливаются самой системой. Другими словами, живые системы автономны. Это не означает, что они изолированы от окружающей их среды. Наоборот, они взаимодействуют с окружением через непрерывный обмен энергией и материей. Но это взаимодействие не определяет их организацию — они остаются самоорганизующимися. Таким образом, автопоэз можно рассматривать как паттерн, лежащий в основе феномена самоорганизации, или автономии; это — важное характерное свойство всех живых систем. Через взаимодействие с окружающей средой живые организмы непрерывно поддерживают и обновляют себя; они используют для этого ресурсы из окружающей среды. Более того, постоянное самосоздание включает также способность формировать новые структуры и новые паттерны поведения. Мы увидим, что создание новизны, приводящее к развитию и эволюции, является глубоким внутренним аспектом автопоэза. Тонкий, но важный момент в определении автопоэза составляет тот факт, что автопоэзная сеть — это не набор отношений между статическими компонентами (каковым, например, является паттерн организации кристалла), но набор отношений между процессами воспроизводства компонентов. Если эти процессы останавливаются, останавливается и вся организация. Другими словами, автопоэзные сети должны непрерывно регенерировать себя, чтобы поддерживать собственную организацию. Это, конечно, хорошо известная особенность жизни. Матурана и Варела видят в различии между взаимоотношениями статических компонентов и взаимоотношениями процессов ключевую разницу между физическими и биологическими феноменами. Поскольку процессы в биологическом феномене включают компоненты, из них всегда можно извлечь описание этих компонентов в чисто физических терминах. Тем не менее, как утверждают авторы, такое чисто физическое описание не охватывает биологический феномен в полной мере. Биологическое объяснение, утверждают они, должно быть описанием взаимоотношений процессов в контексте автопоэза. Диссипативная структура — структура живых систем Описывая паттерн жизни как автопоэзную сеть, Матурана и Варела делают основной акцент на организационной закрытости этого паттерна. Когда структуру живой системы описывает Илья Пригожин, он, наоборот, уделяет главное внимание открытости этой структуры потоку энергии и материи. Таким образом, живая система как открыта, так и закрыта — она открыта структурно, но закрыта организационно. Через систему непрерывно протекает поток материи, но она поддерживает устойчивую форму и обеспечивает это автономно посредством самоорганизации. Чтобы подчеркнуть это кажущееся парадоксальным сосуществование изменений и устойчивости, Пригожин ввел термин «диссипативные структуры». Я уже упоминал, что не все диссипативные структуры являются живыми системами, и, чтобы наглядно показать сосуществование непрерывного потока и структурной устойчивости, удобнее обратиться к простым, неживым диссипативным структурам. Одна из простейших структур такого типа — завихрение в потоке воды, например, водоворот в сливном отверстии ванны. Вода непрерывно проходит сквозь водоворот, и все же его характерная форма, хорошо известные спирали и сужающаяся воронка остаются замечательно устойчивыми (рис. 7–4). Это — диссипативная структура. Более близкое рассмотрение источника и прохождения такого водоворота вскрывает ряд достаточно сложных феноменов". Представьте себе ванну с неглубокой и неподвижной водой. Когда сток открывается, вода начинает вытекать, образуя радиальный поток в направлении стока и ускоряясь под влиянием гравитационной силы по мере приближения к сливному отверстию. Таким образом, устанавливается плавный, единый поток. Однако плавное состояние потока удерживается недолго.
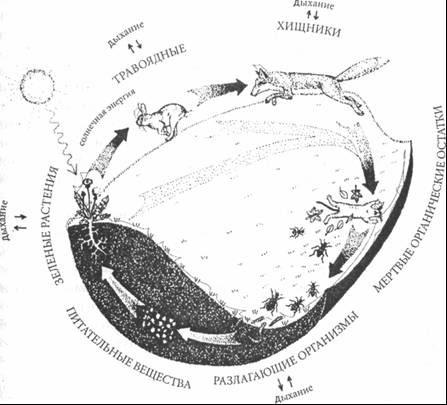
Рис. 7–4 Воронка при сливе воды в ванной Мелкие нерегулярности в движении воды, движении воздуха над поверхностью воды и возмущения в трубе стока приведут к тому, что с одной стороны стока окажется немного больше воды, чем с другой, и тогда в потоке появляется вихревой, круговой компонент движения. По мере того как частицы воды движутся вниз в направлении стока, их радиальная и круговая скорости нарастают. Радиально они ускоряются под действием силы гравитации, а скорость вращения возрастает оттого, что уменьшается радиус вращения: так фигуристка ускоряет обороты, прижимая руки к телу при выполнении пируэта12. В результате частицы воды движутся вниз по спиральным траекториям, образуя сужающуюся трубку линий потока, известную как воронка. Так как основной поток все еще радиален и направлен к центру, воронка непрерывно сдавливается под напором воды со всех сторон. Это давление уменьшает ее радиус и еще больше ускоряет вращение. Используя язык Пригожина, можно сказать, что вращение вносит неустойчивость в изначально однородный поток. Сила тяготения, давление воды и постоянно уменьшающийся радиус воронки — все это, вместе взятое, непрерывно ускоряет вихревое движение жидкости. Это беспрерывное ускорение завершается, однако, не катастрофой, а новым устойчивым состоянием. По достижении определенной скорости вращения в игру вступают центробежные силы: они отталкивают воду от стока по радиусу. Как результат, на изначально плоской поверхности воды над стоком образуется углубление, которое быстро превращается в воронку. В конце концов внутри водоворота формируется миниатюрный воздушный торнадо, а на водной поверхности воронки возникают достаточно сложные нелинейные структуры — барашки, волны и завихрения. Через некоторое время сила тяготения, влекущая воду вниз в направлении стока, давление воды, направленное внутрь потока, и центробежные силы, расталкивающие поток в стороны, уравновешивают друг друга; устанавливается устойчивое состояние, в котором тяготение поддерживает поток энергии высокого уровня, а трение рассеивает некоторую небольшую ее часть. Действующие силы теперь взаимосвязаны через самобалансирующиеся петли обратной связи, которые обеспечивают устойчивость структуре водоворота в целом. Подобные высокоустойчивые диссипативные структуры образуются иногда во время грозы при особых атмосферных условиях. Ураганы и торнадо представляют собой вихри бешено вращающегося воздуха; они могут перемещаться на огромные расстояния и высвобождать разрушительные силы, не проявляя значительных изменений в структуре своего вихря. Подробности процессов в этих атмосферных вихрях гораздо богаче, чем в случае воронки воды в ванной, поскольку здесь появляется несколько новых факторов — разница температур, расширение и сжатие воздуха, эффекты влажности, конденсация и испарение и т. п. Соответственно, гораздо более сложными и разнообразными, чем в водоворотах, оказываются структуры воздушных вихрей и режимы их поведения. Грозы могут превращаться в диссипативные структуры характерных размеров и форм; при особых условиях некоторые из них даже разделяются на два отдельных урагана. Метафорически мы можем представить себе и живую клетку как некий вихрь, т. е. устойчивую структуру, которую постоянно пронизывает поток материи и энергии. Но силы и процессы, действующие в клетке, совершенно другие и гораздо более сложные, чем в вихре. Если балансирующие силы в вихре имеют механический характер, причем доминирует сила тяготения, то соответствующие силы в клетке — химической природы. Точнее говоря, именно каталитические петли в автопоэзной сети клетки действуют как самобалансирующиеся петли обратной связи. Подобным же образом, источник неустойчивости в водовороте носит механический характер и возникает как следствие начального вращательного импульса, а в клетке существуют различные типы неустойчивости, и их природа — химическая, а не механическая. Они тоже берут начало в каталитических циклах, составляющих главную особенность всякого метаболического процесса. Важнейшим свойством этих циклов является то, что они действуют не только как самобалансирующие, но и как самоусиливающие петли обратной связи, способные толкать систему все дальше и дальше от равновесия, пока она не достигнет порога устойчивости. Этот порог называется тонкой бифуркации, или точкой неустойчивости; в таких точках могут спонтанно возникать новые формы порядка, полагая начало развитию и эволюции. Математически точка бифуркации представляет резкое изменение траектории системы в фазовом пространстве13. Внезапно может появиться новый аттрактор — и поведение всей системы идет в новом направлении. Тщательное изучение Пригожиным точек бифуркации выявило еще некоторые замечательные свойства диссипативных структур, о чем пойдет речь в следующей главе14. Диссипативные структуры, формируемые водоворотами или ураганами, могут поддерживать свою устойчивость лишь до тех пор, пока через структуру проходит устойчивый поток материи из окружающей среды. Точно так же, живая диссипативная структура, например организм, нуждается в постоянном проходящем сквозь систему потоке воздуха, воды и пищи из окружающей среды, чтобы оставаться живой и поддерживать свой порядок. Обширная сеть метаболических процессов поддерживает систему в далеком от равновесия состоянии и, через содержащиеся в ней петли обратной связи, вызывает бифуркации, обеспечивая тем самым развитие и эволюцию. Обучение — процесс жизни Три ключевых критерия жизни — паттерн, структура и процесс — так неразрывно переплетены, что трудно обсуждать их раздельно; вместе с тем, необходимо понимать и различия между ними. Автопоэз, паттерн жизни, — это набор взаимоотношений между процессами производства; а диссипативная структура может быть понята только в контексте метаболических и эволюционных процессов. Таким образом, процессуальное измерение присуще критерию как паттерна, так и структуры. В зарождающейся теории живых систем процесс жизни — как непрерывное воплощение автопоэзного паттерна организации в диссипативной структуре — идентифицируется с обучением, процессом познания. Это предполагает радикально новую концепцию разума, которая, возможно, является самым революционным и волнующим аспектом этой теории, поскольку обещает наконец полностью преодолеть картезианское разделение разума и материи. Согласно теории живых систем, разум — это не вещь, а процесс, сам процесс жизни. Другими словами, организационная деятельность живых систем на всех уровнях жизни — это деятельность умственная. Взаимодействие живого организма — растения, животного или человека — с окружающей его средой есть взаимодействие познавательное, или ментальное. Так жизнь и познание становятся неразрывно связанными. Разум — или, более точно, ментальный процесс — имманентен материи на всех уровнях жизни. Новая концепция разума была разработана, независимо друг от друга, Грегори Бэйтсоном и Умберто Матураной в 60-е годы. Бэйтсон, постоянный участник конференций Мэйси в ранние годы кибернетики, стал пионером в применении системного мышления и кибернетических принципов в нескольких новых областях15. В частности, он разработал системный подход к душевным заболеваниям и кибернетическую модель алкоголизма, а в результате пришел к определению ментального процесса как системного феномена, характерного для живых организмов. Бэйтсон сформулировал ряд критериев, которым удовлетворяет система, обладающая разумом16. Любая система, отвечающая этим критериям, способна развивать процессы, которые мы ассоциируем с разумом, — обучение, память, принятие решений и т. п. По Бэйтсону, разум — это необходимое и неизбежное следствие определенной сложности, возникающей задолго до того, как в организме формируется мозг и центральная нервная система. Он также подчеркивал, что разум свойствен не только индивидуальным организмам, но также социальным и экологическим системам. Впервые Бэйтсон представил свою новую концепцию ментального процесса в 1969 году в докладе на конференции по душевному здоровью, проходившей на Гавайях17. В том же году Матурана представил другую формулировку той же основной идеи на конференции по обучению, организованной Хайнцом фон Форстером в Чикаго18. Так два ученых, находившихся под сильным влиянием кибернетики, одновременно пришли к одной революционной концепции разума. Тем не менее их методы радикально различались, как и их язык, на котором они описывали свое новаторское изобретение. Бэйтсон мыслил преимущественно категориями паттернов и отношений. Его основная цель, как и цель Матураны, состояла в том, чтобы найти паттерн организации, общий для всех живых существ. «Какой паттерн, — спрашивал он, — связывает краба с омаром, орхидею с примулой и всех их со мной? И меня с тобой?»19 Бэйтсон считал, что для точного описания природы нужно попытаться говорить на языке природы, а это и есть, как он подчеркивал, язык взаимоотношений. Взаимоотношения, по Бэйтсону, это самая сущность живого мира. Биологическая форма складывается из отношений, а не из частей; Бейтсон настаивал, что это же относится и к способу человеческого мышления. Поэтому он назвал книгу, в которой изложил свою концепцию ментального процесса, «Разум и природа: необходимое единство». Бэйтсон обладал уникальной способностью улавливать тончайшие природные феномены в состоянии сосредоточенного наблюдения. Это не было обычное научное наблюдение. Каким-то образом он умел наблюдать растение или животное всем своим существом, с симпатией и страстью. И когда он говорил о растении, он описывал его с любовью и с мельчайшими деталями, используя язык, которым, по его убеждению, сама природа говорит об общих принципах — а он извлекает их из своего непосредственного контакта с растением. Его трогала красота, проявляемая в сложности паттернов природных взаимоотношений, и описание этих паттернов доставляло ему настоящее эстетическое наслаждение. Бэйтсон разработал критерии ментального процесса интуитивно, лишь на основе своего пристального наблюдения над живым миром. Ему было очевидно, что феномен разума неразрывно связан с феноменом жизни. Всматриваясь в живой мир, он видел, что организационная деятельность этого мира по своему существу ментальна. По его собственным словам, «разум — это суть живого бытия»20. Несмотря на ясное понимание единства разума и жизни — или разума и природы, как он выражался, — Бэйтсон никогда не спрашивал, «что есть жизнь». Он никогда не ощущал потребности в разработке теорий или моделей живой системы, которые составили бы концептуальную основу для его критериев ментального процесса. Разработка именно такой основы была научной целью Матураны. По совпадению — или по интуиции? — Матурана одновременно бился над двумя вопросами, которые, как ему казалось, толкают его в противоположных направлениях: «Какова природа жизни?» и «В чем суть обучения?»21. В конце концов он обнаружил, что ответ на первый вопрос — автопоэз — обеспечивает ему теоретическую основу для ответа на второй. Результатом явилась системная теория обучения, разработанная Матураной и Варелой; иногда ее называют теорией Сантьяго. Главное положение теории Сантьяго, как и теории Бэйтсона, — тождество обучения (процесса познания) с процессом жизни22. Это положение радикально расширяет традиционную концепцию разума. По теории Сантьяго, для существования разума мозг отнюдь не необходим. У бактерии или растения нет мозга, но есть разум. Простейшие организмы способны к восприятию и, следовательно, к обучению. Они не видят, но тем не менее воспринимают перемены в окружающей среде — различие между светом и тенью, жарой и холодом, высокой и низкой концентрацией некоторых химических веществ и т. п. Таким образом, новое понимание обучения, или процесса познания, гораздо шире, чем понятие мышления. В него входят восприятие, эмоции и деятельность — весь процесс Жизни. В мире людей обучение также включает язык, понятийное мышление и все другие атрибуты человеческого сознания. Общее понятие, однако, гораздо шире и может даже не включать мышление. Теория Сантьяго, по моему мнению, обеспечивает первое последовательное научное мировоззрение, действительно преодолевающее картезианский раскол. Разум и материя более не представляются двумя изолированными категориями, но рассматриваются как различные аспекты, различные измерения единого феномена Жизни. Чтобы проиллюстрировать концептуальный прогресс, выраженный этим единым взглядом на Разум, Материю и Жизнь, следует вернуться к вопросу, который более ста лет смущал ученых и философов. Как взаимоотносятся между собой Разум и мозг? Нейробиологи еще в XIX веке знали, что структуры мозга и ментальные функции тесно связаны между собой, однако подробности взаимоотношений между Разумом и мозгом всегда оставались тайной. Еще в 1994 году издатели антологии «Сознание в философии и когнитивная нейробиология» честно признавались в предисловии: «Хотя все и согласны с тем, что Разум имеет некоторое отношение к мозгу, все еще не существует общего согласия по поводу конкретной природы этой взаимосвязи»23. В теории Сантьяго взаимоотношения между разумом и мозгом просты и ясны. Наконец-то отброшена декартовская характеристика разума как мыслящей вещи (rescogitans). Разум — не вещь, а процесс, процесс обучения, тождественный процессу Жизни. Мозг — специфическая структура, посредством которой разум осуществляет свою деятельность. Взаимосвязь между Разумом и мозгом, таким образом, представляет собой взаимосвязь между процессом и структурой. Мозг, конечно, не единственная структура, с помощью которой осуществляется процесс обучения. Вся диссипативная структура организма участвует в процессе обучения, независимо от того, обладает ли организм мозгом и центральной нервной системой. Более того, недавние исследования убедительно показали, что в организме человека нервная, иммунная и эндокринная системы, которые традиционно рассматривались как три изолированные системы, фактически формируют единую когнитивную сеть24. Новый синтез Разума, Материи и Жизни, который будет подробно рассмотрен в последующих главах книги, включает два концептуальных обобщения. Взаимозависимость паттерна и структуры позволяет объединить два подхода к пониманию Природы, которые были раздельными и конкурировали на протяжении всей истории западной науки и философии. Взаимозависимость процесса и структуры позволяет ликвидировать разрыв между Разумом и Материей, который тормозил науку со времен Декарта. Взятые вместе, эти два обобщения обеспечивают три взаимозависимых концептуальных измерения нового научного понимания Жизни. Примечания к главе 7 MaturanaandVarela (1987), p. 47. Вместо «паттерна организации» авторы просто используют термин «организация». См. выше, с. 34–35. См. выше, с. 112 и далее. См. выше, с. 103 и далее. См. выше, с. 103–106. См. выше, с. 99— 100. 7. Maturana and Varela (1980), p. 49. 8. См. Сарга(1982),р. 119. 9. См. ниже, с. 263. Чтобы осуществлять это, ферменты используют другую, дополнительную цепочку ДНК в качестве шаблона для заменяемой секции. Таким образом, двойная цепочка ДНК весьма существенна для этих восстановительных процессов. Я благодарен Вильяму Холлоуэю за исследовательскую поддержку в работе над феноменом водоворота. Говоря техническим языком, этот эффект является следствием сохранения углового момента. См. выше, с. 154–155. См. ниже, с. 208–209. См. выше, с. 72–73. Бэйтсон сначала опубликовал обсуждение этих критериев, изначально названных «ментальными характеристиками»; его можно найти в двух эссе, «TheCyberneticsofSelf: ATheoryofAlcoholism» и «PathologiesofEpistemology», оба напечатаны в Bateson (1972). Более детальное обсуждение см. в Bateson (1979), p. 89ff. Более подробное обсуждение бэйтсоновских критериев ментального процесса см. ниже, Приложение, с. #305 и далее. См. Bateson (1972), р. 478. См. выше, с. 113–114. Bateson (1979), р. 8. Цитируется по Сарга (1988), р. 88. См. выше, с. 112–114. См. ниже, с. 285 и далее. Revonsuo and Kamppinen (1994), p. 5. См. ниже, с.302 и далее. Глава 8 Диссипативные структуры Структура и изменение С самых ранних дней становления биологии философы и ученые заметили, что живые формы самыми на первый взгляд загадочными способами сочетают устойчивость структуры с гибкостью изменений. Как вихри, они зависят от постоянного потока материи, проходящего сквозь них; как пламя, они преобразуют материалы, которыми питаются, чтобы поддерживать свою деятельность и расти; но, помимо всего этого и в отличие от вихря и пламени, живые структуры совершенствуются, размножаются и эволюционируют. Еще в 40-е годы Людвиг фон Берталанфи назвал живые структуры открытыми системами, чтобы подчеркнуть их зависимость от непрерывных потоков энергии и ресурсов. Он ввел термин Fliessgleichgewicht(«текучее равновесие»), чтобы отразить сосуществование равновесия и потока, структуры и изменения — во всех формах жизни1. Теперь экологи изображают экосистемы в виде схем потоков, отмечая пути прохождения энергии и материи в различных пищевых сетях. Такие исследования показывают, что круговая переработка является ключевым принципом экологии. Будучи открытыми системами, все организмы в экосистеме производят отходы, но то, что является отходами для одного вида, служит пищей для другого, поэтому все отходы непрерывно перерабатываются и экосистема в целом, в самом общем итоге, существует без отходов. Зеленые растения играют жизненно важную роль в потоке энергии, пронизывающем все экологические циклы. Корни выбирают из земли воду и минеральные соли, которые в виде соков поднимаются к листьям и там соединяются с углекислым газом (СО2), поступающим из воздуха; так образуются сахара и другие органические соединения (в их число входит и целлюлоза — главный структурный элемент стенок клетки). В ходе этого чудесного процесса, известного как фотосинтез, солнечная энергия преобразуется в химическую и связывается в органических веществах, в то время как кислород освобождается и снова поступает в воздух, откуда его потребляют другие растения и животные в процессе дыхания. Соединяя воду и минералы с солнечным светом и СО2, зеленые растения тем самым связывают землю и небо. Мы привыкли считать, что деревья и травы вырастают из земли, но на самом деле большая часть их вещества происходит из воздуха. Основной объем целлюлозы и других органических соединений, образующихся в процессе фотосинтеза, состоит из тяжелых атомов углерода и кислорода; именно эти элементы растения забирают прямо из воздуха в форме СО2. Таким образом, вес полена почти целиком «набран» из воздуха. Когда полено сгорает в камине, кислород и углерод опять соединяются в СО2 и мы получаем — в виде света и тепла — часть солнечной энергии, которая была затрачена на производство дерева. На рис. 8–1 изображена схема типичного пищевого цикла. По мере того как растения поедаются животными, которых, в свою очередь, поедают другие животные, питательные вещества растений проходят по пищевым сетям, а энергия рассеивается в виде тепла через дыхание и выделения. Отходы, а также мертвые животные и растения перерабатываются так называемыми «разлагающими организмами» (насекомыми и бактериями): в ходе этой переработки из отходов освобождаются первоначальные (базовые) питательные вещества и их снова поглощают зеленые растения. Таким образом, питательные вещества и другие основные элементы непрерывно циркулируют по всей экосистеме, причем энергия рассеивается на каждой стадии. Так осуществляется афоризм Юджина Одума: «Материя циркулирует, энергия рассеивается»2. Единственным отходом экосистемы в целом оказывается тепловая энергия дыхания: она рассеивается в атмосфере и непрерывно пополняется через фотосинтез за счет солнечного излучения. Наша иллюстрация, конечно, сильно упрощена. Реальные пищевые циклы могут быть поняты только в контексте гораздо более сложных пищевых паутин, в которых первоначальные, базовые питательные элементы представлены многими химическими соединениями. В последние годы наши знания в области пищевых паутин значительно расширились и усовершенствовались благодаря Гайя- теории, которая показывает сложное переплетение живых и неживых систем во всей биосфере — растений и камней, зверей и атмосферных газов, микроорганизмов и океанов.
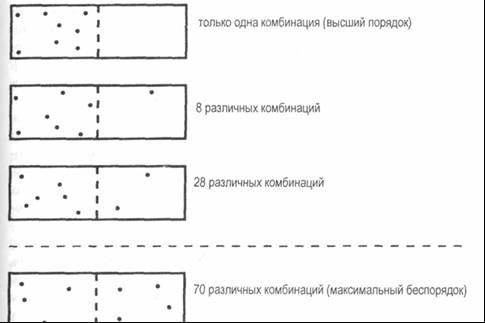
Рис. 8–1. Типичный пищевой цикл Более того, поток питательных веществ через организмы экосистемы не всегда однороден и гладок, но часто сопровождается импульсами, перепадами и разливами. По словам Пригожина и Стенгерс, «энергетический поток, который пересекает [организм], чем-то напоминает реку, которая большей частью течет спокойно, но время от времени устремляется вниз водопадом, высвобождая часть содержащейся в ней энергии»3. Понимание живых структур как открытых систем было важным новым подходом, который, однако, не решил загадку сосуществования структуры и изменения, порядка и рассеяния, пока Илья Пригожий не сформулировал свою теорию диссипативных структур4. Как Берталанфи объединил понятия потока и равновесия для описания открытых систем, так и Пригожий объединил «диссипацию» (рассеяние) и «структуру», чтобы выразить две кажущиеся противоречивыми тенденции, которые сосуществуют во всех живых системах. Однако концепция диссипативных структур Пригожина идет гораздо дальше теории открытых систем, поскольку включает также представление о точках неустойчивости, в которых могут возникать новые структуры и новые формы порядка. Теория Пригожина связывает главные характеристики живых форм в последовательную концептуальную и математическую модель, которая предполагает радикальный пересмотр многих фундаментальных идей, касающихся структуры, — переносит акцент от устойчивости к неустойчивости, от порядка к неупорядоченности, от равновесия к неравновесным состояниям, от бытия к становлению. В центре мировоззрения Пригожина лежит сосуществование структуры и изменения, «покоя и движения»; он изящно поясняет это ссылкой на древнюю скульптуру: Каждый великий период науки предполагал некоторую модель природы. Для классической науки это были часы; для XIX века, периода Промышленной Революции, это был глохнущий мотор. Какой же символ изберем мы? Наше разумение может быть выражено ссылкой на скульптуру — от индейского, доколумбового искусства до наших времен. В самых прекрасных произведениях скульптуры, будь то танцующий Шива или миниатюрные храмы Герреро, отчетливо проявляется стремление соединить покой с движением, время остановленное с временем уходящим. Мы убеждены, что это противоречие подарит нашему времени свою неповторимость5. Неравновесные состояния и нелинейность Ключ к пониманию диссипативных структур лежит в осознании того, что они поддерживают себя в устойчивом состоянии, далеком от равновесия. Эта ситуация настолько отличается от феномена, описываемого классической наукой, что мы сталкиваемся с трудностями традиционного языка. Словарные определения понятия «устойчивый» включают «фиксированный», «не колеблющийся» и «неизменный» — все они неадекватно описывают диссипативные структуры. Живой организм характеризуется непрерывным потоком и изменениями в обмене веществ, включающем тысячи химических реакций. Химическое и тепловое равновесие наступает тогда, когда все эти процессы прекращаются. Другими словами, организм в состоянии равновесия — это мертвый организм. Живые организмы непрерывно поддерживают себя в далеком от равновесия состоянии, которое, по сути, есть состояние жизни. Сильно отличаясь от равновесия, это состояние, тем не менее, сохраняет устойчивость в течение продолжительных периодов времени, что означает, как и в случае вихря, что поддерживается одна общая структура, несмотря на непрекращающийся поток и изменение компонентов. Пригожий понял, что классическая термодинамика — первая наука, трактующая сложные системы, — не подходит для описания далеких от равновесия систем из-за линейной природы ее математической структуры. Близко к состоянию равновесия — в диапазоне классической термодинамики — находятся процессы типа потока, однако они слабы. Система всегда развивается в сторону стационарного состояния, в котором генерация энтропии (или беспорядка) сведена к минимуму. Другими словами, система минимизирует свои потоки, функционируя предельно близко к состоянию равновесия. В этом диапазоне потоковые процессы могут быть описаны линейными уравнениями. Чем дальше от равновесия, тем потоки становятся сильнее, увеличивается выработка энтропии, и тогда система больше не стремится к равновесию. Наоборот, здесь уже могут встретиться неустойчивости, ведущие к новым формам порядка, которые отодвигают систему все дальше и дальше от состояния равновесия. Другими словами, вдали от равновесия диссипативные структуры могут развиваться в формы все более возрастающей сложности. Пригожин подчеркивает, что характеристики диссипативной структуры не могут быть выведены из свойств ее частей, но обусловлены «сверхмолекулярной организацией»6. Корреляции дальнего типа проявляются как раз в точке перехода от равновесия к неравновесному состоянию, и, начиная с этого момента, система ведет себя как единое целое. Вдали от равновесия потоковые процессы в системе взаимосвязаны через многочисленные петли обратной связи, а соответствующие математические уравнения нелинейны. Чем дальше диссипативная структура от равновесия, тем выше степень сложности и нелинейности описывающих ее математических уравнений. Учитывая критическую связь между неравновесным состоянием и нелинейностью, Пригожий и его коллеги разработали нелинейную термодинамику для далеких от равновесия систем, использовав для этого аппарат теории динамических систем — новую математику сложных систем, которая тогда только начинала развиваться7. Линейные уравнения классической термодинамики, как отмечал Пригожий, можно анализировать с помощью точечных аттракторов. Какими бы ни были начальные условия системы, она «увлекается» к стационарному состоянию с минимальной энтропией, предельно близко к равновесию, и ее поведение полностью предсказуемо. Как выражается Пригожий, системы в линейном диапазоне «склонны забывать свои начальные условия»8. За пределами линейного диапазона ситуация совершенно другая. Нелинейные уравнения, как правило, имеют больше чем одно решение; чем выше степень нелинейности, тем больше решений. Это означает, что новые ситуации могут возникать в любой момент. Говоря математическим языком, система в этом случае попадает в точку бифуркации, где может отклониться в совершенно другое состояние. Далее мы увидим, что поведение системы в точке бифуркации (т. е. по какому из нескольких возможных направлений она пойдет) зависит от предыдущей истории системы. В нелинейном диапазоне начальные условия уже «не забываются». Кроме того, теория Пригожина показывает, что поведение далекой от равновесия диссипативной структуры не подчиняется ни одному из универсальных законов: оно уникально для данной системы. Вблизи точки равновесия мы находим повторяющиеся феномены и универсальные законы. По мере удаления от равновесия, мы движемся от универсального к уникальному, в направлении богатства и разнообразия. Это, конечно, хорошо известная характеристика жизни. Наличие точек бифуркации, в которых система может пойти по любому из нескольких различных направлений, предполагает, что неопределенность является еще одной характеристикой теории Пригожина. В точке бифуркации система может сделать «выбор» — этот термин здесь используется метафорически — между несколькими возможными направлениями, или состояниями. Какое направление она выберет, будет зависеть от истории системы и различных внешних условий и никогда не может быть предсказано. В каждой точке бифуркации существует неустранимый элемент случайности. Неопределенность в точках бифуркации представляет собой один из двух типов непредсказуемости в теории диссипативных структур. Другой тип, характерный также для теории хаоса, обусловлен высокой степенью нелинейности уравнений и проявляется даже тогда, когда бифуркации отсутствуют. Из-за многократных петель обратной связи — или, математически, многократных итераций — мельчайшая погрешность в вычислениях, вызванная практической необходимостью определенного округления цифр, неизбежно значительно повышает степень неопределенности, делая предсказания невозможными9. Как неопределенность в точках бифуркации, так и неопределенность «хаотического типа» из-за повторяющихся итераций предполагают, что поведение диссипативной структуры может быть предсказано лишь на короткий промежуток времени. После этого системная траектория ускользает от нас. Таким образом, теория Пригожина, как квантовая теория и теория хаоса, еще раз напоминает нам, что научное знание обеспечивает не более чем «ограниченное окно во вселенную»10. Стрела времени По Пригожину, признание неопределенности как ключевой характеристики естественных феноменов является частью серьезного пересмотра научной концептуальности. Тесно связан с этим концептуальный сдвиг и в научных представлениях о необратимости и времени. В механистической парадигме ньютоновской науки мир рассматривался как полностью причинный и детерминированный. Все, что происходило, имело определенную причину и приводило к определенному следствию. Будущее любой части системы, равно как и ее прошлое, в принципе, могло быть рассчитано с абсолютной определенностью, если состояние этой системы в любой данный момент времени известно во всех подробностях. Этот строгий детерминизм нашел свое самое яркое выражение в знаменитых словах Пьера-Симона Лапласа: Интеллект, который в данное мгновение знает все силы, действующие в природе, и положение всех вещей, из которых состоит мир, — буде сей интеллект достаточно обширен, дабы подвергнуть эти данные анализу, — единой формулой охватит движения громадных тел во вселенной и мельчайшие передвижения атомов; ничто не вызовет у него сомнения, и будущее, равно как и прошлое, предстанет его взору11. В этом лапласианском детерминизме не делается различия между прошлым и будущим. И то и другое заложено в настоящем состоянии мира и в ньютоновых уравнениях движения. Все процессы здесь строго обратимы. Будущее и прошлое чередуются, здесь нет места истории, новаторству или творчеству. Необратимые эффекты (например, трение) отмечались в классической ньютоновской физике, но ими всегда пренебрегали. В XIX столетии ситуация изменилась решительным образом. С изобретением тепловых двигателей необратимость рассеяния энергии при трении, вязкость (сопротивление жидкости течению) и тепловые потери оказались в центре внимания новой науки термодинамики, которая выдвинула идею стрелы времени. В это же время геологи, биологи, философы и поэты начали размышлять над изменением, ростом, развитием и эволюцией. Философия XIX столетия глубоко интересовалась природой становления. В классической термодинамике необратимость, при всей своей важности как понятия, всегда ассоциировалась с рассеянием энергии и потерями. Пригожий фундаментально изменил такой подход в своей теории диссипативных структур, показав, что в живых системах, функционирующих вдали от равновесия, необратимые процессы играют конструктивную и важную роль. Химические реакции — базовые процессы жизни — являются примером необратимых процессов. В ньютоновском мире не может быть ни химии, ни жизни. Теория Пригожина показывает, как каталитические петли — особого типа химические процессы, исключительно важные для живых организмов12, — приводят к состояниям неустойчивости через многократную усиливающую обратную связь и как в последовательных точках бифуркации возникают структуры постоянно нарастающей сложности. «Необратимость, — заключает Пригожий, — есть механизм извлечения порядка из хаоса»13. Таким образом, концептуальный сдвиг в науке, предложенный Пригожиным, означает переход от детерминированных, обратимых процессов к неопределенным, необратимым. Поскольку необратимые процессы играют значительную роль в химии и жизни, при всем том что взаимозаменяемость будущего и прошлого является неотъемлемой частью физики, похоже, что пригожинский пересмотр концепций должен рассматриваться в более широком контексте — том самом, который обсуждался в начале этой книги в связи с глубокой экологией как часть сдвига научной парадигмы от физики к наукам о жизни14. Порядок и беспорядок Стрела времени, как она представляется в классической термодинамике, не указывает на возрастающий порядок, она направлена в противоположную сторону. Согласно второму закону термодинамики, физические феномены проявляют тенденцию к движению от порядка к беспорядку, в сторону непрерывно возрастающей энтропии15. Одно из величайших достижений Пригожина состоит в разрешении парадокса двух противоречивых взглядов на эволюцию — физического и биологического: один представляет идею глохнущего мотора, другой описывает мир, эволюционирующий в сторону возрастающего порядка и сложности. По словам самого Пригожина, «Вот вопрос, преследующий нас более ста лет: какое значение имеет эволюция живого существа в мире, описанном термодинамикой, т. е. в мире непрерывно нарастающего беспорядка?»16 По теории Пригожина, второй закон термодинамики все еще верен, но взаимосвязь между энтропией и беспорядком уже видится в новом свете. Чтобы усвоить это новое представление, нам следует рассмотреть классические определения энтропии и порядка. Концепция энтропии как меры рассеяния энергии на тепло и трение была представлена в XIX веке Рудольфом Клаузиусом, немецким физиком и математиком. Клаузиус определил энтропию, создаваемую в тепловом процессе, как рассеянную энергию, деленную на температуру, при которой происходит процесс. Согласно второму закону термодинамики, энтропия нарастает, по мере того как продолжается тепловой процесс; рассеянная энергия никогда не может быть восстановлена, а направление в сторону непрерывно нарастающей энтропии определяет стрелу времени. Хотя рассеяние энергии на тепло и трение — общеизвестное и привычное явление, сразу же после формулировки второго закона возник интригующий вопрос: что конкретно вызывает эту необратимость? В ньютоновской физике эффектами трения, как правило, пренебрегали, считая их не слишком существенными. Тем не менее эти эффекты можно учитывать и в ньютоновской системе. В принципе, утверждали ученые, можно использовать ньютоновы законы движения для описания рассеяния энергии на молекулярном уровне в форме каскадов столкновений. Каждое из этих столкновений — обратимое событие, поэтому нет ничего невозможного в том, чтобы запустить этот процесс в обратном направлении. Тогда получается, что рассеяние энергии, необратимое на макроскопическом уровне и отвечающее второму закону и обычному опыту, состоит из полностью обратимых событий на микроскопическом уровне. Где же здесь, в таком случае, вкрадывается необратимость? В начале века эта тайна была разгадана австрийским физиком Людвигом Больцманом, одним из великих теоретиков классической термодинамики. Больцман вложил новый смысл в понятие энтропии и установил связь между энтропией и порядком. Следуя рассуждениям основателя статистической механики Джеймса Кларка Максвелла17, Больцман предложил простой мысленный эксперимент, позволяющий исследовать энтропию на молекулярном уровне18. Представьте, что у нас есть коробка, рассуждал Больцман, разделенная на два равных отсека воображаемой перегородкой в центре, и восемь различных молекул, пронумерованных от единицы до восьми подобно бильярдным шарам. Сколько существует способов такого распределения этих частиц в коробке, чтобы их определенное количество находилось по левую сторону перегородки, а остальные — по правую? Для начала поместим все восемь частиц в левый отсек. Это можно сделать лишь одним способом. Если же мы решим поместить семь частиц налево, а одну — направо, то получим восемь способов, так как единственной частицей в правом отсеке может быть любая из восьми частиц. Поскольку молекулы различны, эти восемь способов представляют собой различные комбинации. Подобным же образом, существует 28 различных комбинаций для шести частиц слева и двух справа. Для всех этих перестановок легко вывести общую формулу19. Из нее следует, что количество способов увеличивается по мере того, как уменьшается разность между числом частиц слева и справа, достигая максимума (70 различных комбинаций) при равном распределении молекул, по четыре на каждой половине (рис. 8–2). Больцман называл различные комбинации комплексиями и связывал их с понятием порядка — чем меньше комплексий, тем выше порядок. Таким образом, в нашем примере первое состояние со всеми восемью частицами на одной стороне отражает самую высшую степень порядка, тогда как равное распределение с четырьмя частицами на каждой стороне представляет максимальный беспорядок.
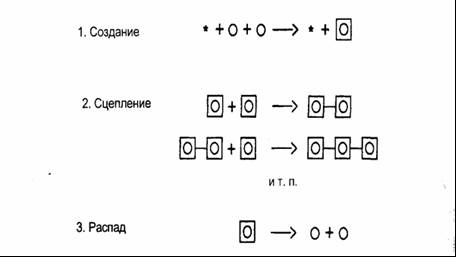
Рис. 8–2. Мысленный эксперимент Больцмана Важно подчеркнуть, что концепция порядка, представленная Больцманом, — это концепция термодинамическая: молекулы находятся в непрерывном движении. В нашем примере перегородка коробки чисто воображаемая, и молекулы в своем беспорядочном движении свободно проходят сквозь нее. В разные моменты времени газ находится в различных состояниях, т. е. количество молекул в отсеках коробки бывает различным; и для каждого из этих состояний число комплексий связано с его степенью порядка. Это термодинамическое определение порядка совершенно отлично от жестких представлений о порядке и равновесии в ньютоновской механике. Рассмотрим другой пример больцмановской концепции порядка, более близкий к нашему повседневному опыту. Представьте, что мы наполняем мешок двумя видами песка — нижнюю половину черным песком, а верхнюю белым. Это состояние высокого порядка; здесь существует лишь одна возможная комплексия. Затем мы встряхиваем мешок, чтобы смешать частицы песка. По мере того как белый и черный песок смешиваются все больше и больше, число возможных комплексий возрастает, а вместе с ней и степень беспорядка, пока мы не получим однородную смесь, состоящую из серого песка, — и максимальный беспорядок. Введя такое определение порядка, Больцман смог анализировать поведение молекул в газе. Используя статистические методы, разработанные Максвеллом для описания беспорядочного движения молекул, Больцман отметил, что число возможных комплексий любого состояния является мерой вероятности того, что газ окажется в этом состоянии. Вот как определяется вероятность. Чем больше комплексий существует для определенной комбинации, тем больше вероятность того, что это состояние установится в газе при беспорядочном движении молекул. Таким образом, число возможных комплексий для определенной комбинации молекул измеряет как степень порядка этого состояния, так и вероятность его установления. Чем выше число комплексий, тем больше беспорядок и выше вероятность того, что газ окажется в этом состоянии. Так Больцман пришел к выводу, что движение от порядка к беспорядку есть движение от менее вероятного состояния к более вероятному. Выражая энтропию и беспорядок через число комплексий, он ввел определение энтропии на языке вероятностных представлений. Согласно Больцману, не существует физического закона, который запрещал бы движение от беспорядка к порядку, но, в силу беспорядочного движения молекул, такое направление весьма маловероятно. Чем больше молекул, тем выше вероятность движения от порядка к беспорядку, а при огромном количестве частиц в газе, эта вероятность практически превращается в определенность. Когда вы трясете мешок с белым и черным песком, вы можете наблюдать, как два типа песчинок разделяются прямо-таки волшебным способом, образуя высокоупорядоченное состояние полного разделения. Но вам, вероятней всего, придется трясти мешок в течение нескольких миллионов лет, чтобы это событие произошло. На языке Больцмана второй закон термодинамики означает, что любая закрытая система стремится к максимально вероятному состоянию, которое представляет собой состояние максимального беспорядка. На математическом языке это состояние может быть определено как аттракторное состояние теплового равновесия. Как только равновесие достигнуто, система, скорее всего, не будет стремиться его покинуть. Временами беспорядочное движение молекул может создавать различные состояния, но они близки к равновесию и существуют лишь в течение коротких периодов времени. Другими словами, система просто флюктуирует (беспорядочно колеблется) вокруг состояния теплового равновесия. Классическая термодинамика, таким образом, пригодна для описания феноменов в состоянии равновесия или близком к равновесию. Пригожинская теория диссипативных структур, напротив, применима к далеким от равновесия термодинамическим феноменам, когда молекулы находятся не в беспорядочном движении, но взаимосвязаны через многочисленные петли обратной связи, описываемые нелинейными уравнениями. В этих уравнениях уже не доминируют точечные аттракторы, а это означает, что система более не стремится к равновесию. Диссипативная структура поддерживает себя вдали от равновесия и может даже уходить все дальше и дальше от него через последовательные бифуркации. В точках бифуркации состояния высшего порядка (в больцмановском смысле) могут возникать спонтанно. Тем не менее это не противоречит второму закону термодинамики. Полная энтропия системы продолжает увеличиваться, но это увеличение энтропии не эквивалентно сплошному увеличению беспорядка. В живом мире порядок и беспорядок всегда создаются одновременно. По Пригожину, диссипативные структуры — это островки порядка в море беспорядка, поддерживающие и даже повышающие свой порядок за счет увеличения беспорядка в окружающей среде. Например, живые организмы забирают упорядоченные структуры (пищу) из окружающей среды, используют их как ресурсы для своих метаболических процессов и рассеивают их как структуры низшего порядка (отходы). Как говорит сам Пригожий, «порядок парит в беспорядке»; при этом общая энтропия продолжает возрастать в соответствии со вторым законом термодинамики20. Это новое представление о порядке и беспорядке радикально переворачивает традиционные научные понятия. В классическом понимании, для которого физика является первоисточником концепций и метафор, порядок эквивалентен равновесию, как, например, в кристаллах и других статических структурах, а беспорядок — неравновесным состояниям, таким как вихри. Новая наука сложных систем, черпающая вдохновение из паутины жизни, показывает, что неравновесное состояние — это источник порядка. Турбулентные потоки воды и воздуха, выглядя хаотическими, на самом деле обладают высокой организацией и сложными паттернами, в которых вихри делятся снова и снова во все более мелких масштабах. В живых системах порядок, возникающий из неравновесных состояний, еще более очевиден; он выражает себя в богатстве, разнообразии и красоте жизни вокруг нас. Во всем живом мире хаос преобразуется в порядок. Точки неустойчивости Точки неустойчивости, в которых происходят непредсказуемые драматические события, где спонтанно возникает порядок и разворачивается скрытая ранее сложность, представляют, вероятно, самый интригующий и замечательный аспект теории диссипативных структур. До Пригожина единственным типом неустойчивости, который изучался более или менее подробно, была турбулентность, вызываемая внутренним трением текущей жидкости или газа21. Леонардо да Винчи провел множество тщательных исследований турбулентных потоков. В XIX веке был поставлен ряд экспериментов, которые показали, что любой поток воды или воздуха становится турбулентным при достаточно высокой скорости — т. е. при достаточно большом «удалении» от равновесия (неподвижного состояния). Исследования Пригожина показали, что для химических реакций это неверно. Химическая неустойчивость не возникает автоматически вдали от равновесия. Для этого необходимы каталитические петли: они подводят систему к точке неустойчивости через многократную усиливающую (положительную) обратную связь22. В этих процессах объединяются два различных феномена — химические реакции и диффузия (физический поток молекул, вызванный разностью концентраций). Соответственно, описывающие их нелинейные уравнения называются уравнениями реакции-диффузии. Они формируют математическую основу теории Пригожина, позволяющую описывать поразительный диапазон типов поведения23. Британский биолог Брайан Гудвин весьма остроумным способом применил пригожинский математический аппарат для моделирования стадий развития весьма специфичной одноклеточной водоросли24. Составив дифференциальные уравнения, которые связывают между собой паттерны концентрации кальция в клеточной жидкости водоросли и механические свойства стенок клетки, Гудвин и его коллеги сумели обнаружить петли обратной связи в процессе самоорганизации, когда в последовательных точках бифуркации появляются структуры нарастающего порядка. Точка бифуркации — это порог устойчивости, где диссипативная структура может либо разрушиться, либо прорваться к одному из нескольких новых состояний порядка. Что на самом деле происходит в этой критической точке, зависит от предыдущей истории системы. В зависимости от того, каким путем она достигла точки неустойчивости, она направится по той или иной ветке после точки бифуркации. Эта важная роль истории диссипативной структуры в критических точках ее развития, обнаруженная Пригожиным даже в простых химических колебаниях, похоже, является физическим началом характерной для всех живых систем связи между структурой и историей. Живая структура, как мы увидим ниже, всегда является записью своего предыдущего развития25. В точке бифуркации диссипативная структура также проявляет исключительную чувствительность к малейшим флюктуациям в окружающей среде. Незначительное случайное отклонение, часто называемое «шумом», может определить выбор направления. Поскольку все живые системы существуют в непрерывно флюктуирующей среде и поскольку невозможно узнать, какое отклонение произойдет в точке бифуркации в «тот самый» момент, мы никогда не можем предсказать будущее направление развития системы. Таким образом, все детерминистские описания оказываются несостоятельными, когда диссипативная структура проходит точку бифуркации. Ничтожные отклонения в окружающей среде предопределяют выбор ветви, по которой эта структура последует. И поскольку в некотором смысле именно эти случайные отклонения приводят к возникновению новых форм порядка, Пригожий ввел описательный термин порядок через флюктуации. Уравнения теории Пригожина — детерминистские уравнения. Они управляют поведением системы на отрезках между точками бифуркации; что касается точек неустойчивости, то здесь решающими оказываются флюктуации — небольшие случайные отклонения. Таким образом, «процессы самоорганизации в далеких от равновесия условиях соответствуют тонкому взаимодействию между случайностью и необходимостью, между флюктуациями и детерминистскими законами»26. Новый диалог с природой Концептуальный сдвиг, предполагаемый теорией Пригожина, включает несколько тесно взаимосвязанных идей. Описание диссипативных структур, которые существуют вдали от равновесия, требует нелинейного математического аппарата, способного моделировать множественные взаимосвязанные циклы обратной связи. В живых организмах, это каталитические циклы (т. е. нелинейные, необратимые химические процессы), которые приводят к точкам неустойчивости через повторяющуюся самоусиливающую обратную связь. Когда диссипативная структура достигает такой точки неустойчивости, называемой точкой бифуркации, в теории появляется элемент неопределенности. В точке бифуркации поведению системы свойственна непредсказуемость. В частности, здесь могут спонтанно возникнуть новые структуры высшего порядка и сложности. Таким образом, самоорганизация, спонтанное возникновение порядка, служит результатом комплексного эффекта неравновесия, необратимости, циклов обратной связи и неустойчивости. Радикальный характер подхода Пригожина очевиден и вытекает из того факта, что к этим фундаментальным идеям редко обращались в традиционной науке, и часто с ними были связаны негативные коннотации. Это следует из самого языка, на котором их описывали. Неравновесный, нелинейность, неустойчивость, неопределенность и т. п. — все это негативные формулировки. Пригожин убежден в том, что этот концептуальный сдвиг, подразумеваемый теорией диссипативных структур, не только критичен для понимания учеными природы жизни, но также помогает нам более полно интегрировать себя в природу. Многие из ключевых характеристик диссипативных структур — чувствительность к малым изменениям в окружающей среде, важность предыдущей истории в критических точках выбора, неопределенность и непредсказуемость будущего — представляются революционными концепциями с точки зрения классической науки, однако служат интегральной частью человеческого опыта. Поскольку диссипативные структуры — это базовые структуры всех живых систем, включая и человеческие существа, это, очевидно, не должно вызывать удивления. Вместо того чтобы быть машиной, природа в целом оказывается более подобной человеку — непредсказуемая, чувствительная к окружающему миру, подверженная влиянию малейших отклонений. Соответственно, адекватный подход к природе с целью изучения ее сложности и красоты состоит не в господстве и контроле, но в уважении, кооперации и диалоге. Действительно, Илья Пригожин и Изабель Стенгерс снабдили свою популярную книгу «Порядок из хаоса» подзаголовком «Новый диалог человека с Природой». В детерминистском мире Ньютона нет места истории и творчеству. В живом мире диссипативных структур история играет важную роль, будущее неопределенно, и эта неопределенность служит основой творчества. «Сегодня, — размышляет Пригожин, — мир, который мы видим снаружи, и мир, который мы ощущаем внутри, сближаются. Это сближение двух миров — вероятно, одно из наиболее важных культурных событий нашего века»27. Примечания к главе 8 1. См. выше, с. 65. 2.Odum(1953). Prigogine and Stengers (1984), p. 156. См. выше, с. 103. Prigogine and Stengers (1984), pp. 22–23. Там же, pp. 143–144. См. выше, с. 131. Prigogine "and Stengers (1984), p. 140. См. выше, с. 144. 10. Prigogine (1989). 11. Цитируется по Сарга (1975), p. 45. Я использовал общий термин «каталитические петли (циклы)» для обозначения множества сложных нелинейных взаимоотношений между катализаторами, включая автокатализ, перекрестный катализ и самоторможение. Более подробно см. PrigogineandStengers (1984), p. 153. Prigogine and Stengers (1984), p. 292. См. выше, с. 28. См. выше, с. 63–64. Prigogine and Stengers (1984), p. 129. См. выше, с. 139–140. См. Prigogine and Stengers (1984), p. 123–124. Если N — общее количество частиц, Ni — частицы на одной стороне, а N2 — на другой, то число различных возможностей определяется формулой Р = N!/N!xN! где N! — факториал N, т. е. 1x2x3… xN. Prigogine (1989). См. Briggs and Peat (1989), p. 45ff. См. Prigogine and Stengers (1984), p. 144ff. Cm. Prigogine (1980), p. 104ff. Goodwin (1994), p. 89ff. См. ниже, с. 238. Prigogine and Stengers (1984), p. 176. Prigogine (1989). Глава 9 Самосозидание Клеточные автоматы Когда Илья Пригожий разрабатывал свою теорию диссипативных структур, он искал простейшие примеры, которые можно было бы описать математически. Он нашел их в каталитических циклах химических колебаний, также известных как «химические часы»1. Это не живые системы, однако те же типы каталитических циклов лежат в основе метаболизма клетки, простейшей из известных живых систем. Поэтому модель Пригожина позволяет нам объяснить существенные структурные особенности клеток на языке диссипативных структур. Умберто Матурана и Франциско Варела следовали подобной стратегии, когда они разрабатывали теорию автопоэза — паттерна организации живых систем2. Они задавали себе вопрос: какое простейшее воплощение автопоэзной сети можно описать математически? Как и Пригожин, они обнаружили, что даже простейшие клетки слишком сложны для математической модели. С другой стороны, они понимали, что поскольку паттерн автопоэза является определяющей характеристикой живой системы, то в природе не найти автопоэзной системы проще, чем клетка. Поэтому, отказавшись от поисков естественной автопоэзной системы, они решили смоделировать ее в виде компьютерной программы. Их подход был аналогичен модели Мира маргариток, разработанной Джеймсом Лавлоком несколькими годами позже3. Однако там, где Лавлока интересовала простейшая математическая модель планеты с биосферой, регулирующей собственную температуру, Матурана и Варела искали простейшую модель сети клеточных процессов, воплощающей автопоэзный паттерн организации. Это означало, что им нужно было разработать особую компьютерную программу: она должна моделировать такую сеть процессов, в которой функция каждого компонента состоит в том, чтобы помогать созданию или трансформации других компонентов сети. Как и в случае клетки, эта автопоэзная сеть также должна создавать собственную границу, которая составляет часть сети процессов, но в то же время определяет ее протяженность. Чтобы найти подходящий математический аппарат для своей задачи, Франциско Варела изучил математические модели самоорганизующихся сетей, разработанные в кибернетике. Двоичные сети, изобретенные Мак-Каллоком и Питтсом в 40-е годы, не обеспечивали достаточного уровня сложности для моделирования автопоэзной сети4; однако оказалось, что более поздние модели сетей — так называемые «клеточные автоматы» — идеально подходят для этой цели. Клеточный автомат представляет собой прямоугольную решетку, состоящую из правильных квадратов, или клеток, — вроде шахматной доски. Каждая клетка может принимать несколько различных «значений», причем существует определенное число соседних клеток, способных влиять на нее. Паттерн, или состояние, всей решетки изменяется дискретно, в соответствии с набором правил перехода, которые вводятся для всех клеток одновременно. Обычно клеточные автоматы полностью детерминированы, но, как мы увидим ниже, в правила легко могут быть включены элементы случайности. Эти математические модели называются автоматами, потому что изначально они были изобретены Джоном фон Нейманном для конструирования машин с возможностью самовоспроизведения. Хотя такие машины так и не были построены, фон Нейманн абстрактно и элегантно показал, что это, в принципе, возможно5. С тех пор молекулярные автоматы широко используются как для имитации природных систем, так и для изобретения большого количества математических игр6. Наверное, самым широко известным примером является игра «Жизнь», в которой каждая клетка может иметь одно из двух «значений», например «черное» или «белое», а последовательность состояний определяется тремя простыми правилами — «рождением», «смертью» и «выживанием». Входе игры возникает поразительное разнообразие паттернов. Некоторые из них «передвигаются»; другие сохраняют стабильность; третьи колеблются или ведут себя еще более сложным образом8. Клеточные автоматы использовались профессиональными математиками и любителями не только для изобретения многочисленных игр; не менее пристально их изучали как математический инструмент для научных моделей. В силу их сетевой структуры и способности работать с большими количествами дискретных переменных, эти математические формы были вскоре признаны и приняты в качестве замечательной альтернативы дифференциальным уравнениям в области имитации сложных систем9. В некотором смысле эти два подхода — дифференциальные уравнения и клеточные автоматы — можно рассматривать как различные математические структуры, соответствующие двум отдельным концептуальным измерениям в теории живых систем — структуре и паттерну. Имитация автопоэзных сетей В начале 70-х Франциско Варела понял, что пошаговые последовательности клеточных автоматов идеальны для компьютерного моделирования и обеспечивают его мощным инструментом имитации автопоэзных сетей. И в 1974 году, совместно с Матураной и ученым-компьютерщиком Рикардо Урибе, Вареле удалось разработать требуемый компьютерный имитатор10. Их клеточный автомат состоит из решетки, в плоскости которой беспорядочно передвигаются «катализатор» и два типа элементов. Они взаимодействуют друг с другом таким образом, что в результате могут образоваться новые элементы обоих видов; одни могут исчезать, а другие связываются друг с другом, образуя цепи. В компьютерных распечатках решетки «катализатор» помечается звездочкой (*). Элемент первого типа, присутствующий в больших количествах, называется «субстратом» и помечается кружком (о); элемент второго типа называется «звеном» и помечается кружком внутри квадрата ([0]). Существует три различных типа взаимодействий и преобразований: два субстрата могуn объединиться в присутствии катализатора, образуя звено; несколько звеньев могут «сцепиться», образуя цепь; любое звено, как свободное, так и входящее в цепь, может распасться снова на два субстрата. В результате некоторого количества преобразований цепь может замкнуться сама на себя. Эти три типа взаимодействия символически изображаются так:
Точные математические предписания (так называемые «алгоритмы»), касающиеся того, когда и как происходят эти процессы, достаточно сложны. Они состоят из многочисленных правил передвижения различных элементов и их взаимодействий". Правила передвижения, например, включают следующие пункты: Субстратам разрешено перемещаться только в незанятые участки(«дырки») решетки; в то же время катализаторам и звеньям разрешено вытеснять субстраты, перемещая их в соседние дырки. Катализатор, кроме того, может вытеснять свободные звенья. Катализатор и звенья могут также меняться местами с субстратами и, таким образом, свободно проходить сквозь их массивы. Субстраты — но не катализатор и не свободные звенья — могут пройти сквозь цепь и занять дырку, расположенную за ней (это имитирует полупроницаемые мембраны клеток). Звенья, связанные в цепь, не могут передвигаться никак. В рамках этих правил фактическое движение элементов и многочисленные подробности их взаимодействия — создание, сцепление и распад — выбираются случайным образом12. Когда запущена имитация на компьютере, генерируется сеть взаимодействий, включающая множество ситуаций случайного выбора, а следовательно, порождающая в свою очередь самые различные последовательности. Авторам удалось показать, что некоторые из этих последовательностей приводят к устойчивым автопоэзным паттернам. Пример такой последовательности взят из их статьи и воспроизведен, в виде семи стадий, на рис. 9–1. В начальном состоянии (стадия 1) одна позиция решетки занята катализатором, а все другие — субстратами. На стадии 2 уже создано несколько звеньев, и, соответственно, теперь в решетке есть несколько дырок. На стадии 3 создано еще больше звеньев и некоторые из них образовали цепи. На стадиях 4–6 производство звеньев и формирование цепей продолжается, и на стадии 7 мы видим, что цепь связанных звеньев замкнулась на себя, охватив катализатор, три звена и два субстрата. Таким образом, цепь сформировала оболочку, проницаемую для субстрата, но не для катализатора. Как только случается такая ситуация, замкнутая цепь может стабилизироваться и превратиться в границу автопоэзной сети. Так случилось и в этой конкретной последовательности. Последующие стадии имитации на компьютере показали, что время от времени некоторые звенья границы могут случайно распадаться, но рано или поздно они заменяются новыми звеньями, созданными внутри оболочки в присутствии катализатора.
Рис. 9–1. Компьютерная имитация автопоэзной сети В ходе длительной имитации цепь и дальше служила оболочкой для катализатора, тогда как звенья продолжали распадаться и заменяться другими. Таким образом, мембраноподобная цепь превратилась в границу сети преобразований, принимая при этом участие в деятельности этой же сети. Другими словами, была смоделирована автопоэзная сеть. Будет ли последовательность такой имитации генерировать автопоэзный паттерн или не будет, в значительной мере зависит от вероятности распада, т. е. от того, насколько часто распадаются звенья. Поскольку тонкое равновесие между распадом и «починкой» основано на случайном движении субстратов сквозь мембрану, случайном создании новых звеньев и случайном перемещении этих звеньев к месту починки, мембрана будет оставаться стабильной только в том случае, если все эти процессы с большой вероятностью завершаются раньше, чем происходит следующий распад. Авторы показали, что при очень маленькой вероятности распада жизнеспособные автопоэзные паттерны действительно могут быть получены13. Двоичные сети Клеточный автомат, разработанный Варелой и его коллегами, стал одним из первых примеров того, как можно моделировать самоорганизующиеся сети живых систем. За последние двадцать лет было изучено множество других имитаций; показано, что эти математические модели способны спонтанно генерировать сложные высокоупорядоченные паттерны, в которых проявляются некоторые важные принципы порядка, наблюдаемые в живых системах. Эти исследования получили новый толчок, когда стало ясно, что недавно разработанные элементы теории динамических систем — аттракторы, фазовые портреты, схемы бифуркации и т. п. — могут быть использованы в качестве эффективных инструментов для анализа моделей математических сетей. Взяв на вооружение эти новые методы, ученые снова обратились к двоичным сетям, разработанным в 40-е годы, и обнаружили, что, хотя это не автопоэзные сети, их анализ приводит к удивительным открытиям в области сетевых паттернов живых систем. Значительную часть этой работы выполнил биолог-эволюционист Стюарт Кауффман совместно с коллегами в институте Санта-Фе, Нью-Мехико14. Поскольку изучение сложных систем с помощью аттракторов и фазовых портретов во многом связано с развитием теории хаоса, перед Кауффманом и его коллегами встал естественный вопрос: какова роль хаоса в живых системах? Мы и теперь еще далеки от полного ответа на этот вопрос, однако работа Кауффмана привела к нескольким интереснейшим идеям. Чтобы понять их, нам придется более пристально рассмотреть двоичные сети. Двоичная сеть состоит из узлов, или переключателей, каждый из которых может находиться в одном из двух состояний, обычно обозначаемых ВКЛ и ВЫКЛ. То есть эта сеть более ограничена в возможностях, чем клеточный автомат, клетки которого могут находиться больше чем в двух состояниях. С другой стороны, узлы двоичной сети не обязательно образуют регулярную решетку, но могут быть соединены между собой более сложными способами. Двоичные сети называют также «булевыми сетями», по имени английского математика Джорджа Буля, который использовал двоичные («да-нет») операции в середине XIX века для разработки символической логики, известной теперь как булева алгебра. На рис. 9–2 показана простая двоичная, или булева, сеть с шестью переключателями, каждый из которых подключен к трем соседним, причем два переключателя находятся в состоянии ВКЛ (черный цвет), а четыре — ВЫКЛ (белый цвет).
Рис. 9–2. Простая двоичная сеть Как и в случае клеточного автомата, паттерн переключателей ВКЛ-ВЫКЛ в двоичной сети меняется дискретным образом. Переключатели соединены между собой так, что состояние каждого переключателя определяется предыдущими состояниями соседних переключателей в соответствии с некоторыми «правилами переключения». Например, для сети, изображенной на рис. 9–2, мы можем выбрать следующее правило: переключатель перейдет в состояние ВКЛ на следующем шаге, если по меньшей мере двое из его соседей на этом шаге будут находиться в состоянии ВКЛ; во всех других случаях А остается в состоянии ВЫКЛ. На рис. 9–3 показаны три последовательности, образовавшиеся по этому правилу. Мы видим, что последовательность А достигает стабильного паттерна, в котором все переключатели находятся в состоянии ВКЛ, через два шага; последовательность В после первого шага колеблется между двумя дополняющими друг друга паттернами; паттерн же С стабилен с самого начала, воспроизводя себя в каждом шаге. Чтобы проанализировать подобные последовательности математически, каждый паттерн, или состояние, сети определяют шестью двоичными (ВКЛ-ВЫКЛ) переменными, т. е. всего двенадцатью переменными. В результате каждого шага система переходит из определенного состояния в определенное последующее состояние, в полном соответствии с правилом переключения.
Рис. 9–3. Три последовательности состояний в двоичной сети Как и в системах, описываемых дифференциальными уравнениями, каждое состояние изображается точкой в 12-мерном фазовом пространстве15. По мере того как, шаг за шагом, сеть переходит из одного состояния в другое, последовательность состояний вычерчивает траекторию в этом фазовом пространстве. Для классификации траекторий различных последовательностей применяется концепция аттракторов. Так, в нашем примере, последовательность А, которая движется к стабильному состоянию, связана с точечным аттрактором, тогда как колеблющееся состояние В соответствует периодическому аттрактору. Кауффман и его коллеги использовали эти двоичные сети для моделирования чрезвычайно сложных систем — химических и биологических сетей с тысячами связанных между собой переменных; такие системы совершенно невозможно описать дифференциальными уравнениями16. Как и в нашем простом примере, последовательность состояний этих сложных систем изображается траекторией в фазовом пространстве. Поскольку число возможных состояний в любой двоичной сети конечно (хотя оно может быть чрезвычайно большим), система должна рано или поздно прийти в то состояние, которое уже встречалось. Когда это произойдет, то следующим шагом система перейдет в то же самое состояние, в которое она переходила и прежде, — поскольку ее поведение полностью детерминировано. Она последовательно повторит тот же цикл состояний. Подобные циклы состояний представляют собой периодические (или циклические) аттракторы двоичной сети. Любая двоичная сеть имеет по крайней мере один аттрактор, но может иметь и больше. Предоставленная самой себе, система в конечном счете закрепится при одном из своих аттракторов и будет в нем оставаться. Периодические аттракторы, вокруг каждого из которых существует своя область притяжения, — наиболее важные математические характеристики двоичных сетей. Обширные исследования показали, что многие живые системы — включая генетические сети, иммунные системы, нейронные сети, системы органов и экосистемы — могут быть представлены в виде двоичной сети, обладающей несколькими альтернативными аттракторами17. Различные циклы состояний в двоичной сети могут значительно различаться по длине. В некоторых сетях они бывают исключительно длинными, и длина эта возрастает по экспоненте с ростом числа переключателей. Кауффман определил аттракторы этих исключительно длинных циклов, насчитывающих миллиарды и миллиарды различных состояний, как «хаотические», поскольку их длина практически бесконечна. Тщательный анализ аттракторов больших двоичных сетей подтвердил то, что кибернетики обнаружили еще в 40-е годы. Некоторые сети хаотичны, поскольку генерируют кажущиеся случайными последовательности и бесконечно длинные аттракторы; другие же генерируют совсем простые аттракторы, соответствующие паттернам высокого порядка. Таким образом, изучение двоичных сетей дает еще одно представление о феномене самоорганизации. Сети, координирующие совместную деятельность тысяч элементов, могут проявлять высокоупорядоченную динамику. У границы хаоса Чтобы установить точную взаимосвязь между порядком и хаосом в этих моделях, Кауффман проверил множество сложных двоичных сетей и разнообразных правил переключения, включая сети, в которых число «входов», или звеньев, различно для разных переключателей. Он обнаружил, что поведение этих сложных паутин можно подытожить, учитывая два параметра: N — число переключателей в сети; К — среднее число входов на каждом переключателе. Для значений К больше 2, то есть в случае множественных взаимосвязей в сети, поведение последней хаотично, но по мере того, как К уменьшается и приближается к 2, устанавливается порядок. Порядок может возникнуть и при более высоких значениях К, если правила переключения «смещены» — например, если ВКЛ преобладает над ВЫКЛ. Подробные исследования перехода от хаоса к порядку показали, что по мере того, как К приближается к 2, двоичные цепи развивают «замороженное ядро» элементов. Это те звенья, которые остаются в одной и той же позиции, ВКЛ или ВЫКЛ, пока система проходит весь цикл состояний. При еще большем приближении К к 2, замороженное ядро создает «стены постоянства», которые вырастают по всей системе, разделяя сеть на отдельные островки меняющихся элементов. Эти островки функционально изолированы. Изменения в поведении одного острова не могут быть переданы сквозь замороженное ядро на другие острова. Если значение К продолжает падать, острова тоже замерзают; периодический аттрактор превращается в точечный, и вся сеть достигает устойчивого, замороженного паттерна. Таким образом, сложным двоичным цепям свойственны три общих режима поведения: упорядоченный режим с замороженными компонентами, хаотический режим без замороженных компонентов и пограничный режим между порядком и хаосом, где замороженные компоненты лишь начинают «таять». Центральная гипотеза Кауффмана заключается в том, что живые системы существуют в этой пограничной области, у края хаоса. Он поясняет, что глубоко в упорядоченном режиме островки деятельности были бы слишком маленькими и изолированными, чтобы сложное поведение могло распространяться по всей системе. Глубоко в хаотическом режиме, с другой стороны, система была бы слишком чувствительна к мельчайшим возмущениям, чтобы поддерживать свою организацию. Таким образом, роль естественного отбора может заключаться в том, чтобы поддерживать живые системы, организованные «на краю хаоса», — потому что здесь они лучше координируют сложное и гибкое поведение, лучше приспосабливаются и развиваются. Чтобы проверить эту гипотезу, Кауффман применил свою модель к генетическим сетям в живых организмах, и ему удалось вывести из нее несколько удивительных и довольно точных предсказаний18. Великие достижения молекулярной биологии, часто именуемые «разгадкой генетического кода», побуждают нас воспринимать цепочки генов в ДНК как некий биохимический компьютер, выполняющий «генетическую программу». Тем не менее последние исследования с нарастающей убедительностью показывают, что этот путь мышления совершенно ошибочен. Фактически он так же неадекватен, как метафора мозга в виде компьютера, обрабатывающего информацию19. Полный набор генов в организме, так называемый «геном», формирует обширную взаимосвязанную сеть, с множеством петель обратной связи, в которой гены прямо и косвенно регулируют деятельность друг друга. По словам Франциско Варелы, «Геном — это не линейный массив независимых генов (проявляющихся как личные качества организма), но в высшей степени взаимно переплетенная сеть множества взаимных воздействий, передаваемых посредством репрессоров и дерепрессоров, экзонов и интроиов, скачущих генов и даже структурных протеинов»20. Когда Стюарт Кауффман начал изучать эту сложную генетическую паутину, он заметил, что на каждый ген в сети напрямую воздействует лишь небольшое число других генов. Более того, уже в 60-е годы было известно, что деятельность генов, как и нейронов, может быть смоделирована на языке двоичных значений ВКЛ-ВЫКЛ. Поэтому, размышлял Кауффман, двоичные сети должны быть подходящими моделями для геномов. Так и оказалось. Действительно, геном моделируется двоичной сетью «на краю хаоса», т. е. сетью с замороженным ядром и изолированными островами «живых», изменяющих свою позицию переключателей. Эта сеть обладает относительно небольшим количеством циклов состояний, представленных в фазовом пространстве отдельными периодическими аттракторами, каждый из которых имеет свою область притяжения. Такая система может подвергаться двум типам возмущений. «Минимальное» возмущение состоит в случайном кратковременном переходе двоичного элемента в противоположное состояние. Оказывается, что каждый цикл состояний модели замечательным образом устойчив к таким минимальным возмущениям. Изменения, вызванные возмущением, не выходят за пределы данного островка деятельности. Другими словами, модель проявляет способность к гомеостазу — свойство, присущее всем живым системам. Другой тип возмущения представляет собой долговременное структурное изменение в сети — например, изменение в паттерне связей или в правилах переключения, — что соответствует мутации в генетической системе. Большинство таких структурных возмущений лишь слегка изменяют поведение сети «на краю хаоса». Некоторые из них, однако, могут сместить траекторию сети в другую сферу притяжения, что приведет к новому циклу состояний и в результате к новому повторяющемуся паттерну поведения. Кауффман видит в этом правдоподобную модель эволюционного приспособления: Сети на границе между порядком и хаосом могут обладать гибкостью быстрой и удачной адаптации через накопление полезных вариантов. В такого рода уравновешенных системах большинство мутаций заканчиваются незначительными последствиями благодаря гомеостатической природе системы. Некоторые мутации, тем не менее, могут вызвать обширные каскады перемен. Поэтому уравновешенные системы, как правило, приспосабливаются к окружающей среде постепенно, однако в некоторых случаях, когда это необходимо, они изменяются быстро21. Еще один ряд впечатляющих особенностей модели Кауффмана касается феномена дифференциации клеток в ходе развития живых организмов. Хорошо известно, что все типы клеток в организме, несмотря на их весьма различные формы и функции, содержат примерно одни и те же генетические инструкции. Считаясь с этим неопровержимым фактом, биологи, занимающиеся проблемами развития, пришли к выводу, что типы клеток различаются не потому, что содержат различные гены, но потому, что в них различны активные гены. Другими словами, структура генетической сети одинакова во всех клетках, однако паттерны генетической деятельности различаются; а поскольку различные паттерны генетической деятельности отвечают различным циклам состояний двоичной сети, Кауффман предположил, что разные типы клеток могут соответствовать разным циклам состояний и, соответственно, разным аттракторам. Эта «аттракторная» модель дифференциации клеток приводит к нескольким интересным предсказаниям22. Каждая клетка человеческого тела содержит около 100 000 генов. В двоичной сети такого размера возможности различных паттернов выражения генов описываются астрономическими цифрами. Тем не менее число аттракторов в такой сети на пороге хаоса примерно равно квадратному корню из числа ее элементов. j Поэтому сеть из 100 000 генов должна выражать себя примерно в 317 ' типах клеток. Это число, выведенное из самых общих положений модели Кауффмана, замечательно приближается к 254 различным типам клеток, обнаруженных в человеческом организме. Кауффман проверил свою аттракторную модель также по числу типов клеток у различных других биологических видов; оказалось, что и эти числа связаны с количеством генов. На рис. 9–4 показаны результаты для нескольких видов23. Очевидно, что количество типов клеток и количество аттракторов соответствующих двоичных цепей возрастает, более или менее параллельно, с увеличением числа генов. Еще два предсказания аттракторной модели Кауффмана касаются стабильности типов клеток. Поскольку замороженное ядро двоичных сетей идентично для всех аттракторов, все клетки организма должны выражать почти один и тот же набор генов и должны различаться по выраженности в небольшом проценте генов. Оказывается, что это действительно так — у всех живых организмов. Аттракторная модель предполагает также, что в процессе развития создаются новые типы клеток — через смещение системы из одной области притяжения в другую. Поскольку у каждой области притяжения есть лишь несколько соседних областей, видоизменение клетки любого типа должно совершаться как переход к немногим непосредственно соседним типам, от них — к следующим соседям и так далее, пока не будет создан полный набор типов клеток. Другими словами, видоизменение Клеток должно происходить в виде последовательно ветвящихся траекторий. Всем биологам известно, что в течение почти 600 миллионов лет дифференциация клеток в многоклеточных организмах происходила именно по этому паттерну. Жизнь в ее минимальной форме Помимо компьютерных имитаций разнообразных самоорганизующихся сетей — как автопоэзных, так и не-автопоэзных, — биологам и химикам позже удалось синтезировать химические автопоэзные системы в лаборатории. Эта возможность была предсказана теоретически Франциско Варелой и Пьером Луиджи Луиси в 1989 году и в дальнейшем реализована в двух типах экспериментов Луиси и его коллегами из Швейцарского политехнического университета (ШПУ) в Цюрихе24. Эти новые концептуальные и экспериментальные достижения резко обострили дискуссию о том, что представляет собой жизнь в ее минимальной форме. Автопоэз, как мы видели, определяется как сетевой паттерн, в котором функция каждого компонента заключается в том, чтобы участвовать в создании или преобразовании других компонентов. Биолог и философ Гэйл Фляйшакер обобщил свойства автопоэзной сети по трем критериям: система должна быть самоограниченной, самопорождающейся и самосохраняющейся25.
Рис. 9–4. Взаимосвязь между количеством генов, типами клеток и аттракторами в соответствующих двоичных сетях для различных биологических видов Самосозидание Самоограничение означает, что протяженность системы определяется границей, которая одновременно является неотъемлемой частью сети. Самопорождение означает, что все компоненты, включая элементы границы, создаются как продукты процессов, происходящих внутри сети. Самосохранение означает, что процессы производства длятся непрерывно таким образом, что все компоненты постоянно заменяются в ходе системных процессов преобразования. Хотя клетка бактерии — простейшая из автопоэзных систем, встречаемых в природе, недавние эксперименты в ШПУ показали, что химические структуры, удовлетворяющие критериям автопоэзной организации, могут быть созданы и в лаборатории. Первая из этих структур, предложенная Луиси и Варелой в их теоретической статье, известна химикам как мицелла. По существу, это капелька воды, окруженная тонким слоем молекул, по форме напоминающих головастиков, с «головками», притягивающими воду, и «хвостами», отталкивающими воду (см. рис. 9–5).
Рис. 9–5. Схематическое изображение капельки-мицеллы При определенных обстоятельствах такая капелька становится вместилищем химических реакций, продуктами которых являются специальные вещества: они самоорганизуются в настоящие пограничные молекулы, которые выстраивают структуру и обеспечивают условия для протекания реакций. Таким образом создается простая химическая автопоэзная система. Как и в компьютерной имитации Варелы, реакции происходят внутри границы, построенной из самих продуктов реакций. После первого примера автопоэзной химии исследователям из ШПУ удалось создать другой тип химической структуры, которая еще больше соответствует клеточным процессам, поскольку ее основные ингредиенты — так называемые жирные кислоты — являются материалом стенок в реальных клетках. Эксперименты состояли в формировании сферических водяных капелек, окруженных оболочками из этих жирных веществ; оболочки имели типичную полупроницаемую структуру биологических мембран (но без их протеиновых компонентов) и генерировали каталитические циклы, приводя к становлению автопоэзной системы. Ученые, проводившие эти эксперименты, предположили, что подобные типы систем могли быть первыми замкнутыми самовоспроизводящимися химическими структурами, возникшими до эволюции бактериальной клетки. Если это верно, то, значит, ученым удалось воспроизвести первые минимальные формы жизни. Организмы и сообщества Исследования по теории автопоэза до сих пор касались, главным образом, минимальных автопоэзных систем — простых клеток, компьютерных имитаций и недавно открытых автопоэзных химических структур. Гораздо меньше исследований было проведено по изучению автопоэза многоклеточных организмов, экологических и социальных систем. Поэтому сегодняшние представления о сетевых паттернах в этих живых системах все еще носят преимущественно умозрительный характер26. Все живые системы — это сети более мелких компонентов, а паутина жизни в целом — многослойная структура живых систем, вложенных в другие живые системы — сети внутри сетей. Организмы — это совокупности автономных, но тесно связанных клеток; популяции — это сети автономных организмов, принадлежащих отдельным видам; а экосистемы — это паутины организмов, как одноклеточных, так и многоклеточных, принадлежащих многим различным видам. Все эти живые системы роднит то, что их мельчайшими живыми компонентами всегда служат клетки, и поэтому мы можем с уверенностью сказать, что все живые системы в конечном счете автопоэзны. Тем не менее возникает интересный вопрос: являются ли более крупные системы, состоящие из автопоэзных клеток, — организмы, сообщества и экосистемы — автопоэзными по своей сути? В книге «Древо познания» Матурана и Варела утверждают, что наше сегодняшнее знание о деталях метаболических направлений в организмах недостаточно для того, чтобы дать ясный ответ, и поэтому они оставляют вопрос открытым: Единственное, что мы можем сказать: [многоклеточные системы] операционно закрыты в своей организации; их идентичность определяется сетью динамических процессов, воздействие которых не выходит за пределы этой сети. Но, наблюдая эту организацию в ее видимой форме, мы ничего не сможем добавить к сказанному27. В дальнейшем авторы подчеркивают, что три типа многоклеточных живых систем — организмов, экосистем и сообществ — радикально различаются по степени автономии своих компонентов. В организмах клеточные компоненты обладают минимальной степенью автономного существования, тогда как компоненты человеческих сообществ, индивидуальные человеческие существа, наделены максимальной степенью автономии, наслаждаясь множеством измерений независимого существования. Сообщества животных и экосистемы занимают промежуточные положения между этими двумя экстремумами. Человеческие сообщества представляют специальный случай из-за решающей роли языка, который Матурана определил как критический феномен в развитии человеческого сознания и культуры28. Если сплоченность социальных насекомых основана на обмене химическими веществами между особями, то социальное единство в человеческих сообществах основано на обмене языковыми сообщениями. Компоненты организма существуют ради функционирования организма, однако человеческие социальные системы существуют также и ради своих компонентов — индивидуальных человеческих существ. Так, по словам Матураны и Варелы: Организм ограничивает индивидуальное творчество своих составляющих, поскольку эти составляющие существуют ради этого организма. Человеческая социальная система усиливает индивидуальное творчество своих компонентов, поскольку она существует ради этих компонентов29. Поэтому организмы и человеческие сообщества — очень разные типы живых систем. Тоталитарные политические режимы часто жестоко ограничивали автономию членов сообщества и, поступая так, деперсонализировали и дегуманизировали их. Фашистские сообщества по режиму своего функционирования ближе к организмам, и поэтому нельзя считать совпадением, что диктаторы так любили использовать метафору общества как живого организма. Автопоэз в социальной сфере Вопрос о том, может ли человеческая социальная система быть описана как автопоэзная, обсуждался довольно широко, и разные авторы предлагали разные ответы30. Главная проблема состоит в том, что автопоэз был точно определен лишь для систем в физическом пространстве и для компьютерных имитаций в математических пространствах. Благодаря «внутреннему миру» понятий, идей и символов, обусловленных человеческой мыслью, сознанием и языком, человеческие социальные системы существуют не только в физическом мире, но также и в символическом социальном мире. Так, человеческая семья может быть описана как биологическая система, обусловленная определенными кровными связями, но также и как концептуальная система, обусловленная определенными ролями и взаимоотношениями, которые могут совпадать — или не совпадать — с кровными связями между ее членами. Эти роли зависят от социальных соглашений и могут значительно меняться в различные периоды времени и в различных культурах. Например, в современной западной культуре роль «отца» может исполнять биологический отец, приемный отец, отчим, дядя или старший брат. Другими словами, эти роли не являются объективными особенностями семейной системы, но служат гибкими и постоянно пересматриваемыми социальными конструктами31. Если поведение в физическом мире управляется причиной и следствием, так называемыми законами природы, то поведение в социальном мире управляется правилами, выработанными социальной системой и часто закодированными в законе. Критическое различие состоит в том, что социальные правила можно нарушить, а законы природы — невозможно. Человеческие существа могут выбирать, подчиняться ли, и в какой форме, социальному правилу; молекулы не могут выбирать, взаимодействовать им или нет32. Учитывая одновременное пребывание социальных систем в двух мирах, физическом и социальном, имеет ли вообще смысл применять к ним понятие автопоэза, а если имеет, то относительно какого мира его следует применять? Оставив вопрос открытым в упомянутой книге, Матурана и Варела впоследствии отдельно выражали свои несколько различные взгляды поэтому вопросу. Матурана рассматривает социальные системы не как автопоэзные образования, но как некую среду, в которой человеческие существа реализуют свой биологический автопоэз через «языкотворчество»33. Варела утверждает, что концепция сети процессов производства, которая лежит в основе определения автопоэза, не может быть применена вне физической сферы, однако для социальных систем может быть определена более широкая концепция организационной закрытости. Эта более широкая концепция сродни автопоэзу, но она не выделяет специально процессов производства34. Автопоэз, по мнению Варелы, можно рассматривать как частный случай организационной закрытости на клеточном уровне и в определенных химических системах. Другие авторы утверждают, что автопоэзная социальная сеть может быть определена, если описание человеческих социальных систем не выходит за рамки социальной сферы. Эта школа мысли была основана в Германии социологом Никласом Люманном, который весьма подробно разработал концепцию социального автопоэза. Суть подхода Люманна состоит в том, чтобы идентифицировать социальные процессы автопоэзной сети как процессы коммуникации: Социальные системы используют коммуникацию как свой особый способ автопоэзного воспроизведения. Их элементами являются сообщения, которые… производятся и воспроизводятся через сеть связи и которые не могут существовать вне такой сети35. Семейная система, к примеру, может быть определена как сеть переговоров, которым присуща кругообразность. Результаты переговоров побуждают к дальнейшим переговорам, и, таким образом, формируются самоусиливающие петли обратной связи. Закрытость сети приводит к общей системе убеждений, объяснений и ценностей — контексту смысла, — которая непрерывно поддерживается дальнейшими переговорами. Коммуникативные акты сети переговоров включают «самопроизводство» и ролей, которыми определяются различные члены семьи, и границ семейной системы. Поскольку все эти процессы происходят в символическом социальном мире, такие границы не могут иметь физическую природу. Это границы ожиданий, конфиденциальности, верности и т. п. Как семейные роли, так и границы непрерывно поддерживаются и переоцениваются посредством автопоэзной сети переговоров. Система Гайи На протяжении последних лет дискуссии по автопоэзу в социальных системах велись очень оживленно; в то же время, как это ни удивительно, по проблеме автопоэза в экосистемах не опубликовано почти ничего. Приходится согласиться с Матураной и Варелой, что множество тенденций и процессов в экосистемах еще не изучены в такой степени, чтобы решать, могут ли экологические сети быть описаны как автопоэзные. Тем не менее было бы безусловно интересно начать дискуссии по автопоэзу с экологами, подобные дискуссиям с социальными исследователями. Прежде всего можно сказать, что функция каждого компонента пищевой паутины состоит в том, чтобы преобразовывать другие компоненты в пределах той же паутины. В то время как деревья извлекают неорганическую материю из окружающей среды, чтобы производить органические соединения, и эти соединения передаются по экосистеме, служа пищей для производства более сложных структур, сеть в целом регулирует себя через множественные петли обратной связи36. Отдельные компоненты пищевой паутины непрерывно умирают, разлагаются и заменяются в ходе собственных процессов преобразования в сети. Достаточно ли этого, чтобы определить экосистему как автопоэзную, покажет время. Кроме прочего, это еще зависит и от ясного понимания границы системы. Перенося наши представления с экосистем на планету в целом, мы сталкиваемся с глобальной сетью процессов производства и преобразования, которая достаточно подробно была описана Джеймсом Лавлоком и Линн Маргулис в их Гайя-теории37. Фактически сегодня можно представить больше свидетельств, подтверждающих автопоэзную природу системы Гайи, чем доказательств существования автопоэза в экосистемах. Планетарная система функционирует в огромных пространственных и временных масштабах. Поэтому конкретно осмысливать Гайю как живое существо весьма затруднительно. Жива ли планета как целое или это относится лишь к ее отдельным частям, и если верно последнее, то к каким частям? Чтобы помочь нам увидеть в Гайе живую систему, Лавлок предложил аналогию дерева37. У растущего дерева лишь очень тонкий слой клеток, расположенных по его периметру, как раз под корой, является живым. Вся древесная масса внутри, более 97 процентоввсего дерева, мертва. Подобным же образом, Земля покрыта тонким слоем живых организмов — биосферой, — которая углубляется в океан на 5–6 миль и поднимается над земной поверхностью примерно на такую же высоту. Итак, живая часть Земли — не что иное, как тонкая пленка вокруг земного шара. Если представить планету в виде мяча, размером с баскетбольный, с нарисованными на нем океанами и странами, то толщина биосферы будет примерно равна толщине краски! Точно так же как корой дерева защищен внутренний тонкий слой живой ткани от повреждений, жизнь на Земле защищена слоем атмосферы, который закрывает нас от ультрафиолетового излучения и других вредных воздействий и поддерживает температуру планеты на уровне, благоприятном для процветания жизни. Ни атмосфера над нами, ни комья земли под нами не являются живыми, но и те, и другие в значительной мере сформированы и преобразованы живыми организмами — точно так же как кора и древесная масса дерева. Открытый космос и недра Земли составляют часть окружающей среды Гайи. Чтобы понять, может ли система Гайи действительно быть описана как автопоэзная сеть, применим три критерия, предложенные Гэйлом Фляйшакером39. Гайя — система определенно самоограниченная, по крайней мере это касается внешней границы, атмосферы. Согласно Гайя-теории, атмосфера Земли создается, преобразуется и поддерживается метаболическими процессами биосферы. Бактерии играют важнейшую роль в этих процессах, влияя на скорость химических реакций, и, таким образом, функционируют как биологический эквивалент ферментов в клетке40. Атмосфера полупроницаема, как клеточная мембрана, и формирует общую часть планетарной сети. Например, она создала защитную теплицу, в которой стало возможным зарождение жизни на планете три миллиарда лет тому назад, хотя в те времена излучение Солнца было на 25 % слабее, чем сейчас41. Система Гайи является также, несомненно, самопорождающейся. Планетарный метаболизм превращает неорганические вещества в органические — в живую материю, — а затем возвращает их в почву, океаны и воздух. Все компоненты сети Гайи, включая элементы атмосферной границы, производятся процессами внутри самой сети. Ключевой характеристикой Гайи является сложное переплетение Живых и неживых систем в пределах единой паутины. Это приводит к возникновению циклов обратной связи совершенно разных масштабов. Циклы горных пород, например, растягиваются на сотни миллионовлет, тогда как организмы, связанные с ними, живут очень недолго. Как метафорически выразился Стивен Хардинг, эколог и соратник Джеймса Лавлока: «Живые существа выходят из камней и уходят в камни»42. И наконец, система Гайи очевидно самосохраняющаяся. Компоненты океанов, почвы и воздуха, равно как и все организмы биосферы, непрерывно заменяются в ходе планетарных процессов производства и преобразования. Похоже, таким образом, что основания для признания Гайи автопоэзной сетью очень вески. И Линн Маргулис, соавтор Гайя-теории, уверенно заявляет: «Мало сомнений в том, что планетарная паутина, включая и нас самих, автопоэзна»43. Убежденность Линн Маргулис в автопоэзности планетарной паутины подкреплена тремя десятилетиями новаторской работы в области микробиологии. Для понимания сложности, разнообразия и самоорганизующих свойств сети Гайи абсолютно необходимо понимание микрокосма — природы, развития, метаболизма и эволюции микроорганизмов. Маргулис внесла огромный вклад в это понимание не только в научном мире. Ей также удалось, в сотрудничестве с Дорион Саган, объяснить свои радикальные открытия ясным и занимательным языком непосвященному читателю44. Жизнь на Земле зародилась примерно 3,5 миллиарда лет назад, и первые два миллиарда лет живой мир состоял исключительно из микроорганизмов. В течение первого миллиарда лет эволюции бактерии — простейшие формы жизни — покрыли планету сложной паутиной метаболических процессов и начали так воздействовать на температуру и химический состав атмосферы, что она стала благоприятной для эволюции высших форм жизни45. Растения, животные и люди поздно появились на Земле: они возникли из микрокосма менее миллиарда лет назад. И даже сегодня известные нам живые организмы функционируют лишь благодаря хорошо развитой связи с бактериальной паутиной жизни. «Мы вовсе не оставили микроорганизмы где-то позади на эволюционной беговой дорожке, — пишет Маргулис. — Мы все окружены ими и состоим из них… [Нам следует] воспринимать самих себя и всю нашу окружающую среду как эволюционную мозаику микрокосмической жизни»46. За всю эволюционную историю жизни более 99 % когда-либо существовавших видов вымерло, однако планетарная паутина бактерий выжила и продолжает регулировать условия для жизни на Земле, как она это делала на протяжении последних трех миллиардов лет. СогласноМаргулис, концепция планетарной автопоэзной сети оправданна, поскольку вся жизнь заключена в самоорганизующуюся паутину бактерий, включающую сложные сети сенсорных и исполнительных систем, которые мы только начинаем познавать. Мириады бактерий, живущих в почве, скалах и океанах, равно как и внутри всех растений, животных и людей, непрерывно регулируют жизнь на Земле: «Именно рост, метаболизм и способность микробов к обмену газами… формируют сложные физические и химические системы с обратной связью, которые, в свою очередь, модулируют биосферу, а вместе с ней и нас, ее обитателей»47. Вселенная в целом Размышляя о планете как о живом существе, невольно задумываешься о системах более крупного масштаба. Не является ли Солнечная система автопоэзной сетью? А Галактика? А что можно сказать о Вселенной в целом? Жива ли Вселенная? Что касается Солнечной системы, то здесь мы с определенной степенью уверенности можем сказать, что она не является живой системой. И именно поразительное различие между Землей и всеми другими планетами Солнечной системы привело Лавлока к формулировке Гайя-гипотезы. В отношении нашей Галактики, или Млечного Пути, мы даже близко не подошли к тем данным, которые могли бы позволить выдвинуть вопрос о том, живая ли это система; а переключая наше внимание на Вселенную в целом, мы тем более упираемся в ограниченность человеческих представлений. Для многих людей, включая меня, в философском и духовном аспекте предпочтительней предположить, что космос в целом жив, нежели думать, что жизнь на Земле существует в окружении безжизненной Вселенной. В рамках науки, однако, мы не можем — или по крайней мере пока не можем — делать подобные заявления. Если мы применим наши научные критерии жизни ко всей Вселенной, мы встретимся с серьезными концептуальными трудностями. Живые системы определяются как открытые непрерывному потоку энергии и материи. Но как мы можем представить себе Вселенную открытой системой, если она, по определению, заключает в себе все сущее? В этом вопросе, похоже, не больше смысла, чем в вопрошании: что происходило до Большого Взрыва? По словам известного астронома сэра Бернарда Лоувелла: Здесь мы приходим к великому барьеру для нашей мысли… Я ощущаю себя так, как будто внезапно въехал в огромную полосу тумана — знакомый мир исчез48. Мы можем сказать относительно Вселенной только то, что потенциал для жизни в избытке существует во всем космосе. Исследования двух последних десятилетий дают достаточно ясную картину геологических и химических особенностей ранней Земли, которые сделали появление жизни возможным. Мы начали понимать, как развивались все более и более сложные химические системы и как они формировали каталитические циклы, которые в конце концов развились в автопоэзные системы49. Изучая Вселенную в целом и, в частности, нашу Галактику, астрономы обнаружили, что характерные химические компоненты, встречающиеся во всех проявлениях жизни, присутствуют там в избытке. Для того чтобы из этих компонентов смогла возникнуть жизнь, требуется тонкий баланс температур, атмосферных давлений и содержания воды. В ходе долгой эволюции Галактики такой баланс с большой вероятностью мог быть достигнут на многих планетах миллиардов планетарных систем, содержащихся в Галактике. Даже в нашей Солнечной системе, как на Венере, так и на Марсе, в период их ранней истории, возможно, существовали океаны, в которых могла зародиться жизнь50. Однако Венера была слишком близка к Солнцу, чтобы обеспечить неторопливую эволюцию. Ее океаны испарились, и в конце концов водород был выбит из молекул воды мощным ультрафиолетовым излучением и исчез в космосе. Мы не знаем, каким образом потерял свою воду Марс, нам лишь известно, что это действительно произошло. Лавлок высказывает по этому поводу догадку, что, возможно, на Марсе существовала жизнь на ранних стадиях его развития, но он потерял ее в результате некоей катастрофы; или же водород пропал там быстрее, чем на ранней Земле, из-за гораздо более слабой силы гравитации на Марсе. Как бы то ни было, похоже, что жизнь на Марсе почти зародилась и что, по всей вероятности, она действительно зародилась и процветает на миллионах других планет по всей Вселенной. Таким образом, хотя в рамках современной науки концепция Вселенной как единой живой системы проблематична, мы уверенно можем сказать, что жизнь с большой вероятностью присутствует в космосе в избытке. Структурное сопряжение Где бы мы ни наблюдали жизнь, от бактерий до широкомасштабных экосистем, мы видим сети с компонентами, которые взаимодействуют друг с другом таким образом, что вся сеть регулирует и организует себя. Поскольку эти компоненты, за исключением элементов клеточных сетей, сами являются живыми системами, реалистичная картина автопоэзных сетей должна включать описание того, как живые системы взаимодействуют между собой и, в общем случае, с окружающей средой. В сущности, такое описание является неотъемлемой частью теории автопоэза, разработанной Матураной и Варелой. Главная особенность автопоэзной системы заключается в том, что она проходит непрерывные структурные изменения, одновременно сохраняя свой паутиноподобный паттерн организации. Компоненты сети непрерывно производят и преобразуют друг друга, и осуществляют они это двумя различимыми способами. Один тип структурных изменений представляют изменения самообновления. Всякий живой организм постоянно обновляет себя, клетки разрушаются и восстанавливаются, ткани и органы заменяют свои клетки в непрерывных циклах. Несмотря на эти непрекращающиеся изменения, организм постоянно поддерживает свою общую идентичность, или паттерн организации. Многие из этих циклических изменений происходят гораздо быстрее, чем это можно представить. Например, наша поджелудочная железа заменяет большинство своих клеток каждые двадцать четыре часа, клетки внутренней оболочки желудка воспроизводятся каждые три дня; наши белые кровяные тельца обновляются за десять дней, а 98 процентов протеина в нашем мозге сменяются меньше чем за месяц. Что еще более поразительно — клетки нашей кожи заменяются со скоростью 100 000 клеток в минуту. Фактически основная часть пыли в наших домах состоит из мертвых клеток кожи. Второй тип структурных изменений в живой системе представляют изменения, посредством которых создаются новые структуры — новые связи в автопоэзной сети. Изменения второго типа — эволюционные, а не циклические; они тоже совершаются непрерывно, либо как последствия влияния окружающей среды, либо как результат внутренней динамики системы. Согласно теории автопоэза, живая система взаимодействует со своей окружающей средой через структурное сопряжение, т. е. через повторяющиеся взаимодействия, каждое из которых запускает структурные изменения в системе. Например, клеточная мембрана непрерывно вводит вещества из своего окружения в метаболические процессы клетки. Нервная система организма изменяет свою внутреннюю связность с каждым сенсорным восприятием. Тем не менее эти живые системы автономны. Окружающая среда лишь запускает структурные перемены, но не определяет и не направляет их51. Структурное сопряжение, как его определяют Матурана и Варела, устанавливает четкое различие между тем, как взаимодействуют со своей окружающей средой живые и неживые системы. Пнуть камень и пнуть собаку — это две совершенно разные истории, как любил отмечать Грегори Бэйтсон. Камень будет реагировать на пинок согласно линейной причинно-следственной цепочке. Его поведение может быть просчитано на основе фундаментальных законов ньютоновской механики. Собака ответит структурными изменениями, согласно своей собственной природе и (нелинейному) паттерну организации. Результирующее поведение, в общем случае, непредсказуемо. Поскольку живой организм отвечает на влияния окружения структурными изменениями, то и эти изменения, в свою очередь, влияют на его последующее поведение. Другими словами, структурно сопряженная система — это обучающаяся система. Пока организм остается живым, он будет структурно сопрягаться со своим окружением. Его непрерывные структурные изменения в ответ на события — и, следовательно, его непрерывное приспособление, обучение и развитие — это и есть ключевые характеристики поведения живых существ. Благодаря его структурному сопряжению, мы называем поведение животного разумным, но мы не применяем этот термин к поведению камня. Развитие и эволюция Продолжая взаимодействовать со своей окружающей средой, живой организм проходит последовательность структурных изменений и со временем формирует свой собственный, индивидуальный путь структурного сопряжения. В каждой точке этого пути структура организма представляет собой запись предыдущих структурных изменений и, следовательно, предыдущих взаимодействий. Живая структура — это всегда запись предыдущего развития, и онтогенез — ход развития индивидуального организма — это история структурных изменений организма. Таким образом, поскольку структура организма в любой точке свое-I го развития представляет запись его предыдущих структурных изменений и поскольку каждое структурное изменение влияет на последующее поведение организма, то из этого следует, что поведение живого организма определяется его структурой. Так, с разных сторон, живая система определяется своим паттерном организации и своей структурой. Паттерн организации определяет своеобразие системы (т. е. ее существенные черты); структура, сформированная последовательностью структурных изменений, определяет поведение системы. По терминологии Матураны, поведение живых систем структурно детерминировано. Эта концепция структурного детерминизма бросает новый свет на старые философские споры о свободе и детерминизме. Согласно Мату-ране, поведение живого организма детерминировано. Однако оно детерминировано не внешними силами, а самой структурой организма — структурой, образовавшейся через последовательность автономных структурных изменений. Получается, что поведение живого организма и детерминировано, и свободно. Больше того, факт структурной детерминированности поведения не означает, что оно предсказуемо. Структура организма просто «обусловливает ход своих взаимодействий и ограничивает структурные изменения, которые могут быть вызваны этими взаимодействиями»52. Например, когда живая система достигает точки бифуркации, как это описано у Пригожина, ее история структурного сопряжения будет определять новые ставшие возможными направления; но по какому направлению пойдет система, остается непредсказуемым. Как и пригожинская теория диссипативных структур, теория автопоэза показывает, что творчество — создание все новых и новых конфигураций — является ключевым свойством всякой живой системы. Особая форма такого творчества — порождение разнообразия через воспроизведение, начиная от простого деления клетки и вплоть до чрезвычайно сложного танца полового размножения. Для большинства живых организмов онтогенез — это не линейный путь развития, но Цикл, и воспроизведение является жизненно важной частью этого Цикла. Миллиарды лет тому назад объединенные способности живых систем к воспроизведению и созданию новизны естественным образом привели к биологической эволюции — творческому раскрытию жизни, которое в виде непрерывного процесса продолжается до сих пор. От самых архаических и простых форм жизни до самых запутанных и сложных современных форм — на этом поле жизнь развернула непрерывный танец, никогда не нарушая основной паттерн своих автопоэзных сетей. Примечания к главе 9 1. См. выше, с. 105. См. выше, с. 114. См. выше, с. 124 и далее. См. выше, с. 100–101. Von Neumann(1966). См. Gardner (1971). В каждом квадрате (3x3) имеется центральная клетка, окруженная 8 соседями. Если три соседние клетки черные, центральная клетка становится черной на следующем шаге («рождение»); если две соседние клетки черные, центральная клетка остается без изменений («выживание»); во всех других случаях клетка становится белой («смерть»). См. Gardner (1970). Великолепный отчет по истории и применению клеточных автоматов см. в Farmer, ToffoliandWolfram (1984), в особенности предисловие Стивена Вольфрама. Более позднее собрание технических статей см. в Gutowitz (1991). Varela, Maturana, and Uribe (1974). Эти передвижения и взаимодействия могут быть формально выражены как математические правила перехода, применяемые одновременно ко всем клеткам. Некоторые из соответствующих математических вероятностей служат переменными параметрами модели. Вероятность распада не должна превышать 0,01 за временной шаг, чтобы вообще могла быть создана жизнеспособная структура, а граница должна со держать не менее 10 звеньев; подробности см. в Varela, MaturanaandUribe (1974). См. Kauffman (1993), pp. 182ff; краткое резюме см. в Kauffman (1991). См. выше, с. 145 и далее. Заметьте, однако, что, поскольку значения двоичных переменных изменяются дискретно, то и их фазовое пространство тоже будет дискретным. См. Kauffman (1993), р. 183. Самосозидание См. там же, p. 191. См. там же, pp. 441 ff. См. выше, с. 83 и далее. Varela et al. (1992), p. 188. Kauffman(1991). 22. См. Kauffman (1993), p. 479. 23.Kauffman(1991). Cm. Luisi and Varela (1989), Bachmann et al. (1990), Walde et al. (1994). Cm. Fleischaker (1990). 26. Недавние дискуссии по вопросам, обсуждаемым ниже, см. в Fleischaker (1992), а также Mingers (1995). Maturana and Varela (1987), p. 89. См. ниже, с. 307 и далее. Maturana and Varela (1987), p. 199. См. Fleischaker (1992); Mingers (1995), p. 119. Mingers (1995), p. 127. Cm. Fleischaker (1992), pp. 131–141; Mingers (1995), pp. 125–126. Maturana (1988); см. также ниже, с. 310–312. Varela (1981). Luhmann(1990). См. выше, с. 121. См. выше, с. 117 и далее. Lovelock (1991), pp. 31 ff. См. выше, с. 227–228. См. выше, с. 110–111. См. Lovelock (1991), pp. 135–136. Harding (1994). См. Margulis and Sagan (1986), p. 66. Margulis (1993); Margulis and Sagan (1986). См. ниже, с. 256 и далее. Margulis and Sagan (1986), pp. 14, 21. Там же, р. 271. Цитируется по Сарга (1975), p. 183. См. ниже, с. 253 и далее. См. Lovelock (1991), р. 127. См. Maturana and Varela (1987), pp. 75ff. Там же, р. 95. Глава 10 Раскрытие жизни О дна из самых замечательных особенностей зарождающейся теории живых систем — необходимо вытекающее из нее новое понимание эволюции. Взгляд на эволюцию как на результат случайных мутаций и естественного отбора сменяется признанием творческого раскрытия Жизни, непрерывно возрастающего разнообразия и сложности — этих неотъемлемых характеристик всякой живой системы. Хотя мутация и естественный отбор по-прежнему признаются важными аспектами биологической эволюции, основное внимание ученых теперь сосредоточено на творчестве, непрерывном стремлении Жизни к обновлению. Чтобы глубже понять фундаментальное различие между старыми и новыми взглядами на эволюцию, рассмотрим кратко историю эволюционной мысли. Дарвинизм и неодарвинизм Первая теория эволюции была сформулирована в начале XIX столетия Жаном Батистом Ламарком, натуралистом-самоучкой, который ввел термин биология и провел обширные исследования в области ботаники и зоологии. Ламарк наблюдал, как животные меняются под воздействием окружающей среды, и полагал, что они могут передавать эти изменения своему потомству. Именно эта передача приобретенных характеристик представлялась ему основным механизмом эволюции. И хотя оказалось, что Ламарк в этом отношении ошибался, его признание феномена эволюции — появления новых биологических форм в истории видов — стало революционным открытием, в значительной степени повлиявшим на последующее развитие этого направления научной мысли. Ламарк оказал сильное влияние на Чарльза Дарвина, который начинал свою научную карьеру как геолог, но во время знаменитой экспедиции на Галапагосские острова заинтересовался биологией. Тщательное изучение фауны острова побудило Дарвина к размышлениям о влиянии географической изоляции на образование видов и привели его в конце концов к формулировке теории эволюции. Дарвин опубликовал теорию в 1859 году в своей монументальной работе «Происхождение видов», а еще через двенадцать лет дополнил ее трудом «Происхождение человека», в котором концепция эволюционной трансформации одних видов в другие расширяется, включая человека. В основу теории Дарвина положены две фундаментальные идеи — случайное отклонение (позже его стали называть случайной мутацией) и естественный отбор. Центральной в этой теории стала догадка, что все живые организмы связаны общим происхождением. Все формы жизни произошли от неких общих предков путем непрерывного процесса отклонений развития в течение миллиардов лет геологической истории. В этом эволюционном процессе производится гораздо больше разновидностей, чем может выжить, поэтому многие особи исчезают в результате естественного отбора; но некоторые варианты выживают и дают жизнь потомкам. В настоящее время эти фундаментальные идеи подробно описаны и подтверждены обширным массивом свидетельств из биологии, биохимии и палеонтологии, и ни один серьезный ученый не подвергает их ни малейшему сомнению. Различия между классической теорией эволюции и зарождающейся новой теорией сосредоточены вокруг динамики эволюции — механизмов, посредством которых осуществляются эволюционные изменения. Собственная концепция Дарвина относительно случайных отклонений базировалась на предположении, весьма характерном для взглядов XIX века на наследственность. Предполагалось, что биологические свойства особи представляют некую «смесь» соответствующих свойств ее родителей, которые вносят в эту смесь более или менее равный вклад. Это означало, что потомок родителя с полезным случайным отклонением унаследует лишь 50 % нового свойства и впоследствии сможет передать только 25 % этого свойства следующему поколению. Таким образом, новое свойство будет быстро затухать, сохраняя ничтожные шансы на сохранение в ходе естественного отбора. Сам Дарвин признавал, что Это серьезный недостаток его теории, который он не может исправить. Интересно, что проблему Дарвина разрешил Грегор Мендель, австрийский монах и ботаник-любитель, и произошло это всего несколько лет спустя после публикации дарвиновской теории. Однако открытие Менделя не было замечено при его жизни и вновь увидело свет лишь в начале XX века, через много лет после его смерти. Основываясь на своих тщательных экспериментах с цветным горохом, Мендель пришел к выводу, что существуют «единицы наследственности» (впоследствии названные генами), которые не смешиваются в процессе воспроизведения, а, напротив, передаются из поколения в поколение, не меняя своей идентичности. Это открытие привело к предположению, что случайные мутации генов не исчезают в течение нескольких поколений, но сохраняются, чтобы в дальнейшем закрепиться — либо исчезнуть полностью — в ходе естественного отбора. Открытие Менделя не только сыграло решающую роль в становлении теории эволюции Дарвина, но и сформировало новое поле исследований — изучение наследственности путем исследования химической и физической природы генов1. Британский биолог Уильям Бэйтсон, страстный приверженец и популяризатор трудов Менделя, в начале века назвал эту новую область генетикой. Между прочим, своего младшего сына он назвал Грегором в честь Менделя. Комбинация дарвиновской идеи постепенных эволюционных изменений с открытой Менделем генетической устойчивостью привела к образованию синтеза, известного как неодарвинизм, который сегодня преподается на биологических факультетах мира как общепризнанная теория эволюции. Согласно неодарвинистской теории, все эволюционные вариации являются следствиями случайных мутаций, т. е. случайных генетических изменений, за которыми следует естественный отбор. Например, если какой-либо вид животных нуждается в густой шерсти, чтобы выжить в холодном климате, он не отвечает на эту потребность отращиванием шерсти, но, вместо этого, развивает все виды случайных генетических изменений, и те особи, чьи изменения вызвали появление густой шерсти, выживают и производят потомство. Таким образом, по словам генетика Жака Моно, «одна лишь случайность лежит в истоках всякого новшества у всех обитателей биосферы»2. По мнению Линн Маргулис, неодарвинизм фундаментально несостоятелен не только потому, что основан на давно устаревших редукционистских понятиях, но и потому, что был сформулирован на неадекватном математическом языке. «Язык жизни — это не просто обычнаяарифметика и алгебра, — утверждает Маргулис, — язык жизни — это химия. Практикующим неодарвинистам не хватает соответствующих знаний, например, в микробиологии, биологии клеток, биохимии… и экологии микробов»3. Одна из причин того, что в наше время ведущие эволюционисты не владеют надлежащим языком для описания эволюционных изменений, по мнению Маргулис, кроется в том, что большинство из них связаны с зоологической традицией и, следовательно, привыкли иметь дело лишь с небольшой, сравнительно недавней частью эволюционной истории. Новейшие исследования в области микробиологии несомненно указывают на то, что главные направления эволюционного творчества сформировались задолго до того, как на сцене появились животные4. Похоже, что центральная проблема неодарвинизма состоит в его редукционистской концепции генома — набора всех генов организма. Великие достижения молекулярной биологии, часто именуемые «разгадкой генетического кода», вылились в тенденцию изображать геном в виде линейной цепи независимых генов, каждый из которых соответствует конкретному биологическому признаку. Однако исследования показали, что отдельный ген может влиять на широкий спектр признаков и, наоборот, часто один лишь признак определяется множеством генов. Таким образом, остается загадкой, как такие сложные структуры, как глаз или цветок, могли развиться путем последовательных мутаций отдельных генов. Настоятельная необходимость изучения координирующей и интегрирующей деятельности всего генома очевидна, однако этому решительно препятствует механистическое мировоззрение, царящее в традиционной биологии. Лишь совсем недавно биологи пришли к пониманию генома живого организма как глубочайшим образом переплетенной сети и начали изучать деятельность этой сети исходя из системной точки зрения5. Системный взгляд на эволюцию Поразительным проявлением генетической целостности стал теперь Уже основательно подтвержденный факт, что эволюция не всегда совершалась в виде непрерывных постепенных изменений, обусловленных Продолжительными цепочками последовательных мутаций. Результаты изучения ископаемых материалов ясно показывают, что на всем протяжении эволюционной истории встречались продолжительные периоды стабильности, или стазиса, не отмеченные генетическими отклонениями, а затем эти периоды сменялись внезапными резкими переходами. Вполне нормальными являются устойчивые периоды протяженностью в сотни тысяч лет. Чтобы не ходить далеко, человеческое эволюционное приключение тоже началось с миллиона лет стабильности первого гоминида, Australopithecusafarensis6. Новая картина эволюции, известная как «пунктирные равновесия», показывает, что внезапные переходы были вызваны механизмами, совершенно отличными от случайных мутаций неодарвинистской теории. Важным аспектом классической теории эволюции является идея о том, что в ходе эволюционных изменений и под давлением естественного отбора организмы постепенно приспосабливаются к окружающей среде, пока не достигнут состояния, достаточно благоприятного для выживания и воспроизведения. В новом системном подходе, наоборот, эволюционные изменения рассматриваются как результат присущей жизни тенденции к созданию нового, причем этот процесс может сопровождаться, но может и не сопровождаться адаптацией к изменяющимся условиям. Соответственно, системные биологи стали изображать геном как самоорганизующуюся сеть, способную к спонтанному производству новых форм порядка. «Мы должны переосмыслить эволюционную биологию, — пишет Стюарт Кауффман. — Большая часть порядка, который мы наблюдаем в организмах, может быть прямым результатом не естественного отбора, но естественного порядка, привилегию работать над которым получил отбор… Эволюция — это не просто "починка на скорую руку"… Это внезапно возникающий порядок, выпестованный и отточенный отбором»7. Всеобъемлющая новая теория эволюции, основанная на недавних открытиях, еще не сформулирована полностью. Однако модели и теории самоорганизующихся систем, о которых шла речь в предыдущих главах этой книги, открывают возможность такой формулировки. Пригожинская теория диссипативных структур показывает, как далекие от равновесия сложные биохимические системы вырабатывают каталитические циклы, приводящие к неустойчивым состояниям и способные производить новые структуры более высокого порядка. Манфред Эйген предположил, что подобные каталитические циклы могли сформироваться еще до появления жизни на Земле, открыв тем самым предбиологическую фазу эволюции. Стюарт Кауффман использовал двоичные сети в качестве математических моделей генетических сетей живых организмов и смог вывести из них несколько известных особенностей видоизменения и эволюции клетки. Умберто Матурана и Франциско Варела описали процесс эволюции в контексте своей теории автопоэза, рассматривая эволюционную историю вида как историю его структурного сопряжения. И, наконец, Джеймс Лавлок и Линн Маргулис в своей Гайя-теории исследовали планетарные измерения раскрытия жизни. Гайя-теория, равно как и ранние работы Линн Маргулис в области микробиологии, выявила несостоятельность узконаправленной дарвинистской концепции приспособления. В реальном живом мире во всей его целостности эволюция не может быть ограничена приспособлением организмов к окружающей среде, поскольку сама эта среда формируется сетью живых систем, способных к приспособлению и творчеству. В таком случае, что же и к чему приспосабливается? Каждый к каждому — это коэволюция. По словам Джеймса Лавлока: Эволюция живых организмов настолько тесно сопряжена с эволюцией окружающей их среды, что вместе они составляют единый эволюционный процесс9. Таким образом, фокус нашего внимания смещается от эволюции к коэволюции — непрерывному танцу, хореография которого обусловлена тонким взаимодействием конкуренции и кооперации, созидания и обоюдного приспособления. Направления творчества Итак, движущую силу эволюции, согласно зарождающейся новой теории, следует искать не в случайных событиях беспорядочных мутаций, но в присущей жизни тенденции к созиданию нового, в спонтанном возникновении нарастающей сложности и порядка. Усвоив суть этого нового понимания, мы можем спросить: в каких же направлениях развивается и выражает себя творчество эволюции? Ответ дает не только молекулярная биология, но и, что еще более важно, микробиология — изучение планетарной паутины мириад Микроорганизмов, которые оставались единственными формами жизни на Земле в течение двух миллиардов лет эволюции. За этот период бактерии непрерывно преобразовывали поверхность и атмосферу Земли и, выполняя эту работу, изобрели все существенные биотехнологии жизни, включая ферментацию, фотосинтез, связывание азота, дыхание и вращательные механизмы для быстрого передвижения. Широкомасштабные исследования в микробиологии в течение последних трех десятилетий определили три основных направления эволюции10. Первое, хотя и наименее важное, представляет собой случайная мутация генов, центральная концепция неодарвинистской теории. Мутация вызывается случайной ошибкой при саморепродуцировании ДНК, когда две цепочки двойной спирали ДНК разъединяются и каждая из них служит шаблоном для построения новой дополнительной цепочки11. Частота возникновения таких случайных ошибок оценивается примерно как одна на несколько сотен миллионов клеток в каждом поколении. Такая частота, похоже, недостаточна для объяснения эволюции огромного разнообразия форм жизни, если учесть тот хорошо известный факт, что большинство мутаций гибельны и лишь очень немногие обусловливают полезные отклонения. Что же касается бактерий, то здесь ситуация несколько иная, поскольку бактерии делятся очень быстро. Они могут делиться примерно каждые двадцать минут, так что, в принципе, из одной менее чем за день может появиться несколько миллиардов отдельных бактерий. Благодаря этой неимоверной скорости воспроизведения, один успешный бактериальный мутант может быстро распространиться в своей окружающей среде, а следовательно, мутации действительно представляют важное эволюционное направление для бактерий. Однако бактерии же развили второе направление эволюционного творчества, притом гораздо более эффективное, чем случайные мутации. Они свободно передают наследственные черты (от одной к другой) в глобальной сети обмена, которая отличается невероятной мощью и эффективностью. Вот как описывают ее Линн Маргулис и Дорион Саган: Последние пятьдесят лет, или около того, ученые наблюдали, как [бактерии] быстро и просто передают различные биты генетического материала другим особям. Каждая бактерия в любой момент времени имеет в своем распоряжении дополнительные гены, иногда попавшие к ней от совершенно других штаммов, для выполнения функций, не предусмотренных в ее собственной ДНК. Некоторые из генетических битов рекомбинируют с собственными генами клетки, другие отправляются дальше… Благодаря этой способности, все бактерии мира в значительной мере обладают доступом к единому резерву генов и следовательно, к адаптивным механизмам всего бактериального царства13. Этот глобальный обмен генами, известный как рекомбинация ДНК, должен занять место среди наиболее поразительных открытий современной биологии. «Если бы генетические свойства микрокосма можно было распространить на более крупные существа, мы бы оказались в научно-фантастическом мире, — пишут Маргулис и Саган, — где зеленые растения делятся генами для фотосинтеза с соседними грибами, а люди могут благоухать или отращивать бивни, занимая гены, соответственно, у розы или моржа»14. Скорость, с которой сопротивляемость лекарствам распространяется среди сообществ бактерий, — вот решающее подтверждение того, что эффективность их коммуникационной сети значительно превосходит эффективность адаптации посредством мутаций. Бактерии могут приспособиться к окружающим условиям в течение нескольких лет там, где более крупным организмам понадобились бы тысячи лет эволюционной адаптации. Таким образом, микробиология преподает нам урок здравого смысла, показывая, что технологии вроде генной инженерии и глобальной коммуникационной сети, которые мы считаем выдающимися достижениями нашей современной цивилизации, используются планетарной паутиной бактерий уже в течение миллиардов лет для регулирования жизни на Земле. Непрерывный обмен генами среди бактерий помимо их основной цепочки ДНК приводит к поразительному разнообразию генетических структур. Это относится и к структуре вирусов, которые не являются автопоэзными системами в полном смысле, но представляют просто цепочки ДНК или РНК в протеиновой оболочке15. По утверждению канадского бактериолога Сорин Сонеа, бактерии, строго говоря, нельзя классифицировать как вид, поскольку все их цепочки могут потенциально разделять одни и те же наследственные черты и, что для них типично, заменять до 15 % своего генетического материала ежедневно. «Бактерия — это не одноклеточный организм, — пишет Сонеа, — это незавершенная клетка… принадлежащая различным химерам, в зависимости от обстоятельств»16. Иначе говоря, все бактерии являются частью единой микрокосмической Паутины Жизни. Эволюция через симбиоз Мутации и рекомбинация ДНК (обмен генами) — вот два основных направления эволюции бактерий. А как же многоклеточные организмы остальных, более крупных форм жизни? Если случайные мутации не служат для них эффективным эволюционным механизмом и если они не обмениваются генами, подобно бактериям, то как же эволюционировали эти высшие формы жизни? Ответ на этот вопрос был дан Линн Маргулис, открывшей третье, совершенно неожиданное направление эволюции. Это направление играет важнейшую роль во всех сферах биологии. Микробиологам хорошо известно, что наиболее фундаментальное разделение всех форм жизни проходит не по линии «растения — животные», как полагает большинство людей, а между двумя типами клеток — обладающими и не обладающими ядром. Бактерии, эти простейшие формы жизни, не имеют клеточных ядер и поэтому называются также прокариотами («безъядерными клетками»), тогда как все другие клетки обладают ядрами и называются эукариотами («ядерными клетками»). Все клетки высших организмов обладают ядром; эукариоты существуют также в виде одноклеточных небактериальных микроорганизмов. Изучая генетику, Маргулис заинтересовалась тем фактом, что в клетке с ядром не все гены находятся именно внутри ядра: Нас всегда учили, что гены расположены в ядре и что ядро является основным управляющим элементом клетки. Еще только изучая генетику, я узнала, что существуют другие генетические системы, с другими паттернами наследственности. С самого начала меня заинтересовали незаконные гены, расположенные вне ядра17. Изучая феномен более подробно, Маргулис выяснила, что все эти «незаконные гены» происходят от бактерий, а затем постепенно пришла к пониманию того, что они принадлежат отдельным живым организмам, маленьким живым клеткам, пребывающим внутри более крупных клеток. Симбиоз, тенденция различных организмов жить в тесной связи друг с другом и часто внутри друг у друга (как бактерии в нашем кишечнике), — широко распространенный и хорошо известный феномен. Однако Маргулис пошла несколько дальше и предложила следующую гипотезу: долговременные формы симбиоза, включая бактерии и другие микроорганизмы, живущие внутри других, более крупных клеток, обусловили и продолжают обусловливать появление новых форм жизни. Маргулис опубликовала свою революционную гипотезу в середине 60-х годов и в течение последующих лет развила ее в зрелую теорию, известную теперь как симбиогенез. Согласно этой теории, создание новых форм жизни через постоянные симбиотические образования рассматривается как основное направление эволюции для всех высших организмов. Наиболее поразительное свидетельство эволюции через симбиоз представляют так называемые митохондрии, «силовые станции» внутри большинства ядерных клеток18. Эти существенные составляющие всех животных и растительных клеток выполняют функции клеточного дыхания; они содержат свой собственный генетический материал и воспроизводятся независимо, в том числе и по времени, от остальной части клетки. Маргулис предполагает, что митохондрии изначально были свободно мигрирующими бактериями, которые в древние времена вторглись в другие микроорганизмы и осели в них на постоянное жительство. «Слившиеся организмы продолжали эволюционировать в более сложные формы жизни, дышащие кислородом, — поясняет Маргулис. — Здесь, таким образом, мы наблюдаем эволюционный механизм более стремительный, чем мутация: симбиотический союз, который становится постоянным»19. Теория симбиогенеза предполагает радикальный сдвиг представлений в эволюционной мысли. В то время как традиционная теория рассматривает раскрытие жизни лишь как процесс расхождения видов, Линн Маргулис утверждает, что образование новых сложных сущностей через симбиоз прежде независимых организмов всегда представляло более мощную и важную эволюционную силу. Этот новый взгляд заставил биологов признать существенную важность кооперации в эволюционном процессе. Если социальные дарвинисты XIX столетия видели в природе лишь конкуренцию — «окровавленные клыки и когти Природы», как выразил это поэт Теннисон, — то мы сейчас начинаем рассматривать непрерывную кооперацию и взаимную зависимость всех форм жизни как центральный аспект эволюции. По словам Маргулис и Саган, «Жизнь взяла верх над планетой не в битве, но постепенно опутав ее сетью»20. Эволюционное раскрытие жизни в ходе миллиардов лет — это история, от которой захватывает дух. Движимая творчеством, присущим всем живым системам, и выраженная в трех отчетливо различных направлениях — мутациях, обмене генами и симбиозе — живая патина планеты распространялась и укреплялась, корректируемая естественным отбором, в виде форм неуклонно нарастающей сложности. Эта история замечательно рассказана Линн Маргулис и Дорион Саган в книге «Микрокосмос»; в значительной степени по материалам их книги написаны последующие страницы21. Нет свидетельств существования какого-то плана, цели или причины в глобальном эволюционном процессе, и, следовательно, нет доказательств прогресса; и все же существуют вполне различимые паттерны развития/Один из них, известный как конвергенция, представляет собой тенденцию организмов к развитию сходных форм для решения сходных проблем, несмотря на различные родовые истории. Так, глаза развивались не один раз — в разные периоды времени и по разным направлениям — у червей, улиток, насекомых и позвоночных. Подобным же образом, крылья независимо эволюционировали у насекомых, рептилий, летучих мышей и птиц. Похоже, что творчество природы не знает пределов. Еще один поразительный паттерн представляют собой повторяющиеся катастрофы — своего рода планетарные точки бифуркации, за которыми следуют интенсивные периоды роста и совершенствования. Так, опасное падение процентного содержания водорода в земной атмосфере более чем два миллиарда лет назад привело к одной из величайших эволюционных инноваций — использованию воды в фотосинтезе. Миллионы лет спустя эта чрезвычайно успешная новая биотехнология породила катастрофический кризис загрязнения — накопление огромных объемов токсичного кислорода. Кислородный кризис, в свою очередь, обусловил эволюцию бактерий, дышащих кислородом: это оказалось еще одним из замечательных нововведений жизни. Позже, 245 миллионов лет назад, вслед за опустошительным, беспрецедентным вымиранием множества видов наступила быстрая эволюция млекопитающих; а 66 миллионов лет назад катастрофа, которая стерла динозавров с лица Земли, расчистила путь для эволюции первых приматов и, наконец, человеческих существ. Эпохи жизни Чтобы отразить схематически процесс раскрытия жизни на Земле, мы используем геологическую шкалу времени, на которой периоды измеряются в миллиардах лет. Процесс начинается с формирования планеты Земля — огненного шара раскаленной лавы — примерно четыре с половиной миллиарда лет назад. Геологи и палеонтологи разбили эти 4,5 миллиарда лет на многочисленные периоды и подпериоды, обозначенные названиями типа «протерозой», «палеозой» или «плейстоцен». К счастью, нам не обязательно помнить все эти технические термины, чтобы представить себе основные стадии эволюции. В эволюции жизни на Земле мы различаем три достаточно объемлющие эпохи, каждая из которых охватывает временной период от одного до двух миллиардов лет и состоит из нескольких отдельных стадий (см. таблицу на стр. 254). Первая эпоха — предбиотическая, в течение которой формировались условия для возникновения жизни. Она длилась один миллиард лет, от формирования Земли до возникновения начальных форм жизни — первых клеток — около 3,5 миллиардов лет назад. Вторая эпоха, длившаяся полных два миллиарда лет, — это эпоха микрокосма, когда бактерии и другие микроорганизмы изобрели все базовые процессы жизни и сформировали глобальные циклы обратной связи для саморегуляции системы Гайи. Около 1,5 миллиардов лет назад были, в основном, сформированы поверхность и атмосфера Земли в их нынешнем виде; микроорганизмы заполнили воздух, воду и почву, циклически перегоняя газы и питательные вещества по своей планетарной сети, как они делают это и сегодня; и, наконец, были созданы условия для перехода к третьей эпохе жизни — макрокосму, — эпохе эволюции более крупных форм жизни, включая и род человеческий. Происхождение жизни В течение первого миллиарда лет после формирования Земли постепенно складывались условия для появления жизни. Изначальный огненный шар был достаточно велик для того, чтобы удерживать атмосферу. Кроме того, он содержал основные химические элементы, из которых предстояло сформироваться строительным блокам жизни. Расстояние от Солнца оказалось оптимальным — достаточно далеким, чтобы начался процесс медленного охлаждения и конденсации, и в то же время достаточно близким, чтобы не наступило сжижение и замерзание газов. После полумиллиарда лет постепенного охлаждения пар, наполнявший атмосферу, наконец сконденсировался; обильные дожди не прекращались тысячелетиями, и на поверхности Земли скопилось столько воды, что из нее образовались неглубокие океаны. В течение этого продолжительного периода углерод — химический костяк жизни — активно соединялся с водородом, кислородом, азотом, серой и фосфором, порождая бесконечное разнообразие химических соединений. Эти шесть элементов — С, Н, О, N, S, Р — и сейчас являются основными химическими ингредиентами всех живых организмов. Эпохи жизни Миллиардов лет назад Стадии эволюции ПРЕДБИОТИЧЕСКАЯ ЭРА формирование условий для жизни 4,5 формирование Земли охлаждение огненного шара раскаленной лавы 4,0 старейшие горные породы конденсация пара 3,8 мелкие океаны соединения на углеродной основе каталитические циклы, мембраны МИКРОКОСМ эволюция микроорганизмов 3,5 первые бактериальные клетки ферментация фотосинтез сенсорные механизмы, движение починка ДНК обмен генами 2,8 тектонические платформы, континенты кислородный фотосинтез 2,5 повсеместное распространение бактерий 2,2 первые ядерные клетки 2,0 закрепление кислорода в атмосфере 1,8 дыхание на основе кислорода1 1,5 формирование поверхности и атмосферы Земли МАКРОКОСМ эволюция более крупных форм жизни 1,2 передвижение 1,0 половое размножение 0,8 митохондрии, хлоропласты 0,7 первые животные 0,6 раковины, скелеты 0,5 первые растения 0,4 сухопутные животные 0,3 динозавры 0,2 млекопитающие 0,1 цветковые растения первые приматы В течение долгих лет ученые обсуждали вероятность возникновения жизни из «химического супа», который настаивался по мере охлаждения планеты и расширения океанов. Было высказано немало гипотез о внезапных событиях, послуживших первичным толчком, — от драматической вспышки мощной молнии и вплоть до осеменения Земли макромолекулами посредством метеоритов. Другие ученые возражали, что вероятность наступления любого из этих событий практически равна нулю. Тем временем, как выяснилось в результате новейших исследований самоорганизующихся систем, нет принципиальной необходимости постулировать какое-либо внезапное событие. Как отмечает Маргулис, «химические вещества соединяются не случайным образом, а упорядочение, по определенным паттернам»22. Окружающая среда ранней Земли благоприятствовала образованию сложных молекул, ставших затем катализаторами для множества химических реакций. Постепенно различные каталитические реакции сомкнулись, образовав сложные каталитические паутины из замкнутых петель: сначала это были просто циклы, затем гиперциклы, затем структуры с сильной тенденцией к самоорганизации и даже самовоспроизведению23. Когда была достигнута эта стадия, определилось и направление предбиологической эволюции. Каталитические циклы эволюционировали в диссипативные структуры и, проходя через последовательные нестабильные состояния (точки бифуркации), образовывали химические системы все большей сложности и разнообразия. В конце концов эти диссипативные структуры начали формировать мембраны — сначала, видимо, из жирных кислот без протеинов, подобно недавно полученным в лаборатории мицеллам24. Маргулис полагает, что именно тогда могли возникнуть многообразные самовоспроизводящиеся химические системы, заключенные в мембрану; некоторое время они эволюционировали и исчезали, прежде чем появились первые клетки: «Должно было развиться множество диссипативных структур, длинных цепочек различных химических реакций, которые эволюционировали, вступали в реакции и разрушались, прежде чем сформировалась и начала с высокой точностью воспроизводиться элегантная двойная спираль нашего древнего предка»25. В этот период, около 3,5 миллиардов лет назад, зародились первые автопоэзные бактериальные клетки и началась эволюция жизни. Как сплеталась бактериальная паутина Существование первых клеток было шатким. Окружающая среда непрерывно менялась, и каждая случайность представляла новую угрозу их выживанию. Перед лицом всех враждебных сил — жесткого облучения солнечным светом, столкновений с метеоритами, наводнений, засух и извержений вулканов — бактериям приходилось захватывать и удерживать энергию, воду и пищу, чтобы оставаться живыми и целыми. Каждый кризис, несомненно, сметал значительную часть первых островков жизни с лица планеты, и это быстро закончилось бы полным уничтожением, если бы не две жизненно важные особенности тех первых форм: бактериальные ДНК способны к точному воспроизведению и осуществляют его с неимоверной скоростью. В силу своего огромного количества бактерии снова и снова творчески реагировали на все угрозы и развивали разнообразные адаптивные стратегии. Так они постепенно распространялись, сначала в водной среде, а затем и в поверхностных слоях осадочных пород и почвы. Очевидно, наиболее важная задача состояла в том, чтобы развить достаточное разнообразие метаболических способов извлечения энергии и пищи из окружающей среды. Одним из первых изобретений бактерий стала ферментация, т. е. расщепление Сахаров и преобразование их в энергетические носители — молекулы АТФ, которые подпитывают энергией все клеточные процессы26. Эта инновация позволила бактериям, способным к ферментации, добывать химические вещества в земле, грязи и воде, защищаясь тем самым и от жесткого солнечного облучения. Некоторые из ферментаторов выработали, помимо этого, способность поглощать азот из воздуха и перерабатывать его в различные органические соединения. Связывание азота, т. е. непосредственный захват его из воздуха, требует огромных затрат энергии, и даже сегодня эта задача под силу лишь немногим специализированным бактериям. Поскольку азот является ингредиентом протеинов во всех клетках, все ныне существующие организмы для своего выживания нуждаются в бактериях, связывающих азот. В самом начале эпохи бактерий фотосинтез — «несомненно самое важное метаболическое усовершенствование в истории жизни на планете»27 — стал первичным резервом жизненной энергии. Первые процессы фотосинтеза, изобретенные бактериями, отличались от тех, что сегодня происходят в растениях. Вместо воды в качестве источника водорода они использовали сероводород — газ, источаемый вулканами. Они соединяли его с солнечным светом и СО2 воздуха, образуя органические соединения, и никогда не вырабатывали кислород. Эти адаптивные стратегии не только позволяли бактериям выживать и развиваться, но и постепенно начали изменять окружающую их среду. Фактически именно бактерии, почти с самого начала своего существования, сформировали первые петли обратной связи, которые в конце концов должны были неминуемо привести к появлению тесно взаимосвязанной системы — жизни и ее окружения. И хотя химия и климат ранней Земли способствовали развитию жизни, это благоприятное состояние не могло бы поддерживаться бесконечно долго без бактериальной регуляции28. По мере того как железо и другие элементы вступали в реакции с водой, высвобождался газообразный водород; он поднимался сквозь атмосферу, где разлагался на атомы. Поскольку эти атомы слишком легки для того, чтобы их удерживало земное тяготение, весь водород должен был улетучиться, учитывая бесконтрольность процесса; через какой-нибудь миллиард лет всем океанам на планете предстояло исчезнуть. К счастью, вмешалась жизнь. На поздних стадиях фотосинтеза стал высвобождаться и поступать в воздух свободный кислород, как это происходит и сегодня, и некоторая его часть соединялась с восходящими потоками газообразного водорода, образуя при этом воду; так сохранялся определенный уровень влажности на планете и предотвращалось испарение океанов. Тем не менее постоянный отбор СО2 из атмосферы в процессе фотосинтеза вызвал другую проблему. В начале эпохи бактерий энергия солнечного излучения была на 25 % меньше, чем сейчас, и СО2 в атмосфере был совершенно необходим, чтобы создавать тепличный эффект и поддерживать температуру планеты в приемлемом диапазоне. Если бы отбор СО2 происходил без какой-либо компенсации, Земля бы замерзла и ранние формы бактерий погибли бы. Эта опасная тенденция была остановлена ферментирующими бактериями, которые, возможно, сформировались еще до появления фотосинтеза. В процессе производства молекул АТФ из Сахаров ферментаторы также вырабатывали метан и СО2 в виде отходов. Последние поступали в атмосферу, где и восстанавливали планетарный тепличный эффект. Таким образом, ферментация и фотосинтез стали взаимно балансирующими процессами системы ранней Гайи. Солнечный свет, проходивший сквозь атмосферу древней Земли, все еще содержал обжигающую ультрафиолетовую радиацию, и теперь бактериям приходилось балансировать между защитой от облучения и необходимостью получать солнечную энергию для фотосинтеза. Это привело к эволюции многочисленных сенсорных систем и двигательных механизмов. Некоторые виды бактерий мигрировали в воды, богатые определенными солями, выполнявшими роль солнечных фильтров; другие нашли защиту в песке; а некоторые тем временем развили пигменты, в которых поглощались вредоносные лучи. Многие виды организовывали огромные колонии — многослойные «скатерти» из микробов, где верхние слои обжигались и умирали, но защищали нижний слой своими мертвыми телами29. Помимо защитной фильтрации, бактерии выработали также механизмы для починки ДНК, поврежденных радиацией, в том числе специально для этого предназначенные ферменты. Сегодня почти все организмы по-прежнему содержат в себе такие «ферменты-ремонтники» — еще одно пережившее миллиарды лет изобретение микрокосмоса30. Вместо того чтобы использовать для починки собственный генетический материал, бактерии иногда заимствовали фрагменты ДНК у своих соседей по густонаселенному окружению. Этот метод постепенно эволюционировал в непрерывный обмен генами, который и определил самое эффективное направление эволюции бактерий. У высших форм жизни рекомбинация генов различных особей связана с воспроизведением, но в мире бактерий два эти феномена протекают независимо. Бактериальные клетки воспроизводятся бесполым путем, но зато они непрерывно обмениваются генами. По словам Маргулис и Саган, Мы обмениваемся генами «вертикально» — через поколения, — тогда как бактерии меняются ими «горизонтально» — непосредственно со своими соседями из того же поколения. В результате получается, что генетически неустойчивые бактерии функционально бессмертны, а для эукариотов пол связан со смертью31. Из-за небольшого числа постоянных генов в бактериальной клетке — как правило, меньше одного процента от числа генов в ядерной клетке — бактерии по необходимости работают командами. Разные виды сотрудничают и помогают друг другу, предоставляя дополнительный генетический материал. Крупные сообщества таких бактериальных команд могут функционировать с согласованностью единого организма, выполняя задачи, которые индивидуально не под силу никакой из них. К концу первого миллиарда лет с момента возникновения жизни Земля кишела бактериями. Были изобретены тысячи биотехнологий — большинство из них, безусловно, известно сегодня, — и, посредством сотрудничества и непрерывного обмена генами, микроорганизмы начали регулировать условия для жизни на всей планете, как они делают это и поныне. Фактически многие виды бактерий ранней эпохи микрокосма дожили, существенно не изменившись, до наших дней. В ходе последующих стадий эволюции, микроорганизмы образовывали союзы и эволюционировали совместно с растениями и животными, и сегодня наша окружающая среда в такой степени переполнена бактериями, что почти невозможно определить, где кончается неодушевленный мир и где начинается жизнь. Мы склонны ассоциировать бактерии с болезнью, но они жизненно важны и для нашего выживания, равно как и для выживания животных и растений. «Если отбросить в сторону наши поверхностные различия, можно сказать, что все мы представляем собой ходячие сообщества бактерий, — пишут Маргулис и Саган. — Весь мир мерцает, как ландшафт пуантилиста, составленный из крошечных живых существ»32. Кислородный кризис Вследствие того, что бактериальная паутина разворачивалась и заполняла все доступные пространства в водах, скалах и грязевых низинах, ее энергетические потребности привели к серьезному водородному истощению атмосферы. Углеводы, играющие существенную роль во всех процессах жизни, представляют собой сложные структуры из атомов углерода, водорода и кислорода. Чтобы построить эти структуры, фотосинтезирующие бактерии извлекали углерод и кислород в виде СО2, подобно современным растениям. Кроме того, они получали водород в форме газа из воздуха и из сероводорода, извергающегося из вулканов. Однако легкий газообразный водород продолжал улетучиваться в космос, и со временем одного сероводорода стало недоставать. Огромное количество водорода, конечно, есть в воде (Н2О), однако связи между молекулами водорода и кислорода в воде гораздо прочнее, чем между двумя атомами водорода в его газе (Н2) или в сероводороде (H2S). Бактерии, осуществляющие фотосинтез, не были способны разорвать эти крепкие связи, пока особый вид сине-зеленых бактерий не изобрел новый тип фотосинтеза, который навсегда решил проблему водорода. Новый эволюционный тип бактерий, предков современных сине-зеленых водорослей, использовал солнечный свет с более высокой энергией (с более короткими длинами волн) для того, чтобы расщеплять молекулы воды на составляющие их водород и кислород. Они забирали водород для формирования Сахаров и других углеводов, а кислород уходил в воздух. Это изъятие водорода из воды, представляющей один из наиболее обильных ресурсов планеты, стало чрезвычайной эволюционной победой, которая очень глубоко повлияла на последующее раскрытие жизни. И Линн Маргулис убеждена в том, что «пришествие кислородного фотосинтеза было тем исключительным событием, которое в конечном итоге привело к формированию нашей современной окружающей среды»33. Благодаря неограниченным запасам водорода, новые бактерии достигли небывалых успехов. Они быстро распространялись по поверхности Земли, покрывая камни и песок сине-зеленой пленкой. И даже сегодня они вездесущи, прорастая в прудах и бассейнах, на влажных стенах и ставнях — везде, где доступен солнечный свет и вода. Однако этот эволюционный успех был оплачен дорого. Как и все быстро распространяющиеся живые системы, сине-зеленые бактерии производили отходы в огромных количествах, и в данном случае отходы оказались крайне токсичными. Это был газообразный кислород — побочный продукт нового типа фотосинтеза на основе воды. Свободный кислород токсичен потому, что он легко вступает в реакции с органическими веществами, производя так называемые свободные радикалы, которые оказывают весьма разрушительное воздействие на углеводы и другие важные биохимические соединения. Так же легко кислород вступает в реакции с атмосферными газами и металлами, вызывая сгорание или коррозию — две наиболее знакомые формы окисления, т. е. соединения вещества с кислородом. Поначалу Земля легко поглощала кислородные отходы. Вулканические и тектонические источники поставляли достаточно металлов и серных соединений, которые быстро связывали свободный кислород, не давая ему закрепиться в воздухе. Однако абсорбируя кислород в течение миллионов лет, связывающие кислород металлы и минералы насытились, и тогда токсичный газ стал накапливаться в атмосфере. Около двух миллиардов лет назад кислородное загрязнение привело к катастрофе в беспрецедентных глобальных масштабах. Многочисленные виды исчезли полностью, и всей бактериальной паутине пришлось фундаментально перестраиваться, чтобы выжить. Было развито множество защитных механизмов и адаптивных стратегий, и, наконец, кислородный кризис привел к одной из величайших и наиболее удачных инноваций во всей истории жизни: Осуществляя один из величайших переворотов всех времен, [сине-зеленые] бактерии изобрели метаболическую систему, которой требовалось то самое вещество, которое представляло собой смертельный яд… Дыхание кислородом — это исключительно эффективный способ отвода и использования реактивности кислорода. Это — идеально контролируемое сгорание, в котором расщепляются органические молекулы и производятся углекислый газ и вода, а в придачу огромное количество энергии… Микрокосм сделал больше, чем просто приспособился: он изобрел работающую на кислороде машину, которая навсегда изменила саму жизнь и ее земную обитель34. С этим замечательным изобретением в распоряжении сине-зеленых бактерий оказались два дополнительных механизма — генерация свободного кислорода через фотосинтез и его поглощение через дыхание. Теперь они могли приступить к формированию петель обратной связи, которые впредь будут регулировать содержание кислорода в атмосфере, поддерживая здесь тонкий баланс, необходимый для развития новых форм, дышащих кислородом35. Содержание свободного кислорода в атмосфере в итоге стабилизировалось на 21 %. Это значение определилось порогом воспламеняемости. Если бы содержание кислорода упало до 15 %, ничто не могло бы гореть. Организмы не смогли бы дышать и погибли бы. Если бы содержание кислорода в воздухе поднялось до 25 %, то сгорело бы все. Возгорание происходило бы спонтанно, и всю планету охватили бы пожары. И Гайя в течение миллионов лет поддерживала атмосферный кислород на уровне, наиболее благоприятном для всех растений и животных. Кроме того, в верхних слоях атмосферы постепенно образовался слой озона (трехатомных молекул кислорода), и с тех пор он защищает жизнь на Земле от жесткого ультрафиолетового излучения Солнца. Так была подготовлена сцена для появления и эволюции более крупных форм жизни — грибов, растений и животных; все это произошло уже в сравнительно короткие сроки времени. Ядерная клетка Первым шагом в направлении высших форм жизни стал симбиоз — новое направление эволюционного творчества. Это случилось около 2,2 миллиардов лет назад и привело к эволюции эукариотических («ядерных») клеток, которые в дальнейшем стали фундаментальными элементами всех растений и животных. Ядерные клетки гораздо крупнее и сложнее, чем бактерии. Если бактериальная клетка содержит единственную цепочку ДНК, свободно плавающую в клеточной жидкости, то ДНК в эукариотической клетке плотно закручена в хромосомы, которые заключены в мембрану внутри клеточного ядра. Количество ДНК в ядерных клетках в сотни раз больше, чем в бактериях. Еще одной поразительной особенностью ядерной клетки является обилие органелл — поглощающих кислород маленьких частиц, которые выполняют ряд исключительно специализированных функций36. Анализ внезапного появления ядерных клеток в истории эволюции, а также открытие органелл как отдельных самовоспроизводящихся организмов привело Линн Маргулис к заключению, что ядерные клетки развились в результате длительного симбиоза — постоянного сосуществования различных бактерий и других микроорганизмов37. Предками митохондрий и других органелл могли быть бактерии-уродцы, которые вторгались в более крупные клетки и воспроизводили себя внутри них. Многие из завоеванных клеток, очевидно, погибали, а вместе с ними и их завоеватели. Однако некоторые хищники не уничтожили своих хозяев, но стали сотрудничать с ними, и в конце концов естественный отбор позволил выжить и эволюционировать лишь организмам, склонным к сотрудничеству. Возможно, клеточные мембраны развились как средство защиты генетического материала клеток-хозяев от нападения завоевателей. За миллионы лет взаимоотношения, основанные на сотрудничестве, стали еще более координированными и тесными, причем органеллы производили потомство, хорошо приспособленное к жизни внутри более крупных клеток, а крупные клетки становились все более зависимыми от своих постояльцев. Со временем бактериальные сообщества стали до такой степени взаимозависимы, что могли функционировать лишь как единые, целостные организмы: Жизнь продвинулась еще на один шаг, от создания сетей свободного генетического обмена к синергии симбиоза. Отдельные организмы сливались воедино, образуя новые целостности, которые представляли собой нечто большее, чем сумма их частей38. Признание симбиоза как главной эволюционной силы имеет важный философский подтекст. Все крупные организмы, включая и нас самих, служат живыми свидетельствами того факта, что деструктивные поведенческие механизмы на большой дистанции несостоятельны. В конце концов агрессоры всегда уничтожают самих себя и расчищают путь для тех, кто знает, как сотрудничать и развиваться. Жизнь в гораздо меньшей степени является конкурентной борьбой за выживание, чем триумфом сотрудничества и творчества. Действительно, со времени создания первых ядерных клеток эволюция шла через все более сложные формы сотрудничества и коэволюции. Эволюционный путь через симбиоз позволил новым формам жизни многократно и всесторонне использовать хорошо опробованные специализированные технологии в разных комбинациях. Например, хотя бактерии получают пищу и энергию, применяя огромное разнообразие остроумных методов, из их метаболических нововведений животными используется только кислородное дыхание — специальная функция митохондрий. Митохондрии присутствуют и в растительных клетках, которые, кроме того, содержат так называемые хлоропласты — зеленые «солнечные станции», ответственные за фотосинтез39. Эти органеллы замечательным образом напоминают сине-зеленые бактерии, которые, по всей вероятности, и были их предками. Маргулис полагает, что проникающие бактерии, как правило, переваривались завоеванными микроорганизмами, но некоторые разновидности, очевидно, сопротивлялись этому перевариванию внутри хозяев40. Они приспосабливались к новому окружению, продолжая вырабатывать энергию через фотосинтез; более крупные клетки вскоре стали зависимы от поступления этой энергии. Обеспечив ядерным клеткам доступ к эффективному использованию солнечного света и кислорода, новые симбиотические взаимоотношения дали им и третье великое эволюционное преимущество — возможность двигаться. Если компоненты бактериальной клетки медленно и пассивно плавают в клеточной жидкости, то составляющие ядерной клетки, похоже, передвигаются более осмысленно; клеточная жидкость течет единым потоком, и вся клетка может ритмично растягиваться или сокращаться или быстро передвигаться как единое целое — что видно на примере кровяных клеток. Как и множество других жизненных процессов, быстрое движение было изобретено бактериями. Самый быстрый член микрокосма — крошечное, напоминающее волосок создание, названное спирохетой («скрученный волос») и известное также как «бактерия-штопор», поскольку двигается по спирали подобно штопору. Прицепляясь симбиотически к более крупным клеткам, подвижная спирохета дает этим клеткам огромное преимущество быстрого перемещения — способности избегать опасности и искать пищу. Со временем бактерии-штопоры утеряли свои индивидуальные черты и эволюционировали в хорошо известные «клеточные кнуты» — flagellae, cilia, и т. п., — которые служат средством перемещения для множества различных ядерных клеток, как бы подстегивая их своими волнообразными движениями. Объединенные преимущества трех типов симбиоза, описанных в предыдущих параграфах, вызвали вспышку эволюционной активности, которая, в свою очередь, породила огромное разнообразие эукариотических клеток. Обладая двумя эффективными способами выработки энергии и радикально возросшей мобильностью, новые симбиотические формы жизни мигрировали в новые окружения, эволюционируя в первые растения и в первых животных, которым в конце концов суждено было покинуть воду и выбраться на сушу. Как научная гипотеза, концепция симбиогенеза — создания новых форм жизни через слияние различных видов — насчитывает едва тридцать лет. Но как культурный миф эта идея, похоже, стара, как само человечество41. Религиозные эпические творения, легенды, волшебные сказки и другие мифические истории всего мира населены фантастическими созданиями — сфинксами, русалками, гриффонами, кентаврами и другими, — появившимися на свет в результате смешения одного или более видов. Как и клетки-эукариоты, эти создания состоят из хорошо знакомых компонентов, но их комбинации непривычны и поразительны. Изображения этих гибридов зачастую ужасны, но многие из них, как это ни забавно, считаются приносящими удачу. Например, бог Ганеша, который обладает человеческим телом с головой слона, — один из наиболее почитаемых в Индии божеств; ему поклоняются как символу удачи и помощнику в преодолении препятствий. Похоже, что каким-то образом коллективному человеческому бессознательному с древнейших времен известно, что продолжительный симбиоз в высшей степени благотворен для всякой жизни. Эволюция растений и животных Эволюция растений и животных за пределы микрокосма осуществлялась через последовательность симбиозов, в которых бактериальные изобретения предыдущих двух миллиардов лет комбинировались в бесконечных проявлениях творчества, пока не были отобраны жизнеспособные формы. Для этого эволюционного процесса характерна возрастающая специализация — от органелл в первых эукариотах до исключительно специализированных клеток у животных. Важным аспектом клеточной специализации является изобретение полового размножения около миллиарда лет тому назад. Мы привыкли думать, что пол и размножение тесно связаны между собой, однако, как отмечает Маргулис, сложный танец полового размножения состоит из нескольких отдельных компонентов, которые развивались независимо и только постепенно обрели взаимосвязь и единство42. Первым компонентом является тип деления клетки, называемый мейозом («уменьшением»), при котором число хромосом в ядре уменьшается ровно наполовину. Так создаются специализированные клетки яйца и спермы. Затем эти клетки трансформируются в процессе оплодотворения, который восстанавливает нормальное число хромосом, и появляется новая клетка — оплодотворенное яйцо. В дальнейшем эта клетка последовательно делится в процессе роста и развития многоклеточного организма. Слияние генетического материала двух разных клеток широко распространено среди бактерий, где оно происходит в виде непрерывного обмена генами, который не связан с размножением. У ранних растений и животных появилась связь между размножением и слиянием генов, которая впоследствии эволюционировала в сложные процессы и ритуалы оплодотворения. Пол был более поздним усовершенствованием. Первые эмбриональные клетки — сперма и яйцо — были почти идентичными, но со временем они эволюционировали в маленькие, быстрые клетки спермы и большие неподвижные яйцеклетки. Связь между оплодотворением и формированием эмбриона образовалась еще позже, в процессе эволюции животных. В мире растений оплодотворение вылилось в сложные паттерны совместной эволюции цветов, насекомых и птиц. По мере того как продолжалась специализация клеток в более крупных и сложных формах жизни, возможности, связанные с самовосстановлением и регенерацией, постепенно снижались. Плоские черви, полипы и морские звезды могут почти полностью регенерировать свои тела из маленьких частиц; ящерицы, саламандры, крабы, омары и многие насекомые все еще способны отращивать потерянные органы или конечности; однако для высших животных регенерация ограничена обновлением тканей в процессе заживания ран. Как последствие этой утери восстановительных функций, все крупные организмы подвержены старению и, в конечном счете, смерти. Тем не менее с половым размножением жизнь изобрела новый тип восстановительного процесса, в котором целые организмы опять и опять формируются заново, с каждым поколением возвращаясь к единичной ядерной клетке. Растения и животные — не единственные многоклеточные создания в живом мире. Как и другие особенности живых организмов, многоклеточность эволюционировала неоднократно, по многим родословным древам жизни, и сегодня все еще существует несколько видов многоклеточных бактерий, а также множество многоклеточных протестов (микроорганизмов с ядерными клетками). Подобно животным и растениям, большинство видов этих многоклеточных организмов формируются последовательным делением клеток, но некоторые из них образуются как объединение клеток от разных, но принадлежащих одному и тому же виду источников. Замечательный пример таких объединений дает слизистая плесень — макроскопический организм, но по своей конституции — протист. Простая слизистая плесень обладает сложным жизненным циклом, включающим подвижную (как у животных) и неподвижную (как у растений) фазу. В животной фазе она зарождается как массив отдельных клеток, которые обычно можно найти в лесу под гниющими бревнами и влажными листьями, где они питаются за счет других микроорганизмов и вянущей растительности. Часто эти клетки едят так много и делятся столь стремительно, что полностью истощают пищевые ресурсы окружающей среды. Когда это происходит, они объединяются в связную массу из тысяч клеток, похожую на слизня и способную ползать по лесной почве, движениями напоминая амебу. Найдя новый источник пищи, плесень вступает в свою растительную фазу, развивая ножку с плодоносной мякотью, очень похожую на гриб. Наконец, плодовая коробочка взрывается, выстреливая наружу тысячи сухих спор, из которых появляются новые отдельные клетки; они теперь будут передвигаться независимо в поисках пищи, начиная новый цикл жизни. Среди разнообразных многоклеточных организаций, которые развились из тесно связанных сообществ микроорганизмов, три — растения, грибы и животные — были столь удачны в отношении размножения, изменчивости и распространения по всей Земле, что биологи классифицировали их как царства — самые широкие категории живых организмов. Всего таких царств пять — бактерии (микроорганизмы без клеточного ядра), протисты (микроорганизмы с ядерными клетками), растения, грибы и животные43. Каждое из царств иерархически делится на подкатегории, или таксоны, начиная с типа и кончая родом и видом. Теория симбиогенеза позволила Линн Маргулис и ее коллегам построить классификацию живых организмов на ясных эволюционных взаимоотношениях. На рис. 10-1 в упрощенной форме показано, как протисты, растения, грибы и животные эволюционировали из бактерий через ряд последовательных симбиозов, подробно описанных ниже. Следуя за эволюцией растений и животных, мы приходим к макрокосму и должны переключить наш временной диапазон с миллиардов лет на миллионы. Самые древние животные развились около 700 млн. лет назад, а первые растения возникли около 200 млн. лет спустя. И те, и другие сначала эволюционировали в воде и вышли на сушу 400–450 млн. лет назад, причем растения опередили животных на несколько миллионов лет. И растения, и животные развили огромные многоклеточные организмы, но если межклеточные связи в растениях минимальны, то клетки животных исключительно специализированы и тесно взаимосвязаны посредством множества сложных звеньев связи. Уровень взаимной координации и управления значительно возрос, когда стали развиваться первые нервные системы; примерно 620 млн. лет назад у животных появились зачатки мозга. Предками растений были волокнистые массы водорослей, обитающие в мелких пронизанных солнцем водах. Время от времени воды высыхали, но некоторым водорослям удалось выжить, размножиться и превратиться в растения. У этих ранних растений, как у сегодняшних мхов, не было ни стволов, ни листьев. Чтобы выжить на суше, им было совершенно необходимо развить стойкие структуры, которые противостояли бы истощению и засухе. Они выполнили эту задачу: они создали лигнин — материал для клеточных стенок, который позволил растениям сформировать крепкие стволы и ветви, а также сосудистые системы для подъема воды от корней к ветвям и листьям.
Рис. 10-1. Эволюционные взаимоотношения между пятью царствами жизни Основной проблемой, возникшей в новом, наземном окружении, был недостаток воды. Творческий ответ растений выразился в том, что они заключили свой зародыш в защитное семя, противостоящее засухе. Теперь они могли ждать со своим развитием до тех пор, пока не окажутся в достаточно влажной среде. Более чем сто миллионов лет, в то время, когда первые сухопутные животные — амфибии — эволюционировали в рептилий и динозавров, буйные тропические заросли семенных папоротников — фактически семенных деревьев, напоминающих гигантские папоротники, — покрывали огромные просторы Земли. Около 200 миллионов лет назад на нескольких континентах появились ледники, и семенные папоротники не смогли пережить долгие холодные зимы. Их сменили вечнозеленые хвойные деревья, похожие на наши современные пихты и ели; высокая холодостойкость позволила им не только пережить зимы, но и завоевать высокогорные области. Еще сто миллионов лет спустя появились цветущие растения, чьи семена были заключены в плоды. С самого начала эти новые цветковые растения эволюционировали совместно с животными, которые с удовольствием поедали их питательные плоды и, в порядке любезности, распространяли непереваренные семена растений. Кооперативные связи продолжали развиваться, и сегодня в них включились люди — садовники, огородники и др., которые не только распространяют семена растений, но и разводят вегетативно бессеменные растения для получения от них плодов. Как замечают Маргулис и Саган, «похоже, что растения весьма сведущи в обольщении нас, животных, заставляя нас делать для них одну из немногих вещей, которая доступна нам, но недоступна им, — передвигаться»44. Завоевание суши Первые животные эволюционировали в воде из сферических и червеобразных масс клеток. Они все еще были слишком малы, но некоторые из них формировали сообщества, которые коллективно строили огромные коралловые рифы в виде плотных кальциевых отложений. Не обладая твердыми частями или внутренними скелетами, ранние животные полностью разлагались после смерти, однако сотню миллионов лет спустя их потомки построили множество изысканных раковин и скелетов, которые оставили отчетливые отпечатки в хорошо сохранившихся ископаемых породах. Для животных адаптация к жизни на суше стала эволюционным подвигом, потребовавшим решительных изменений в системе органов. Серьезнейшую проблему в условиях недостатка воды, конечно, представляло обезвоживание; хватало, однако, и других проблем. В атмосфере было неизмеримо больше кислорода, чем в океанах, что требовало других органов для дыхания; были необходимы различные типы кожи для защиты от нефильтрованного солнечного облучения; требовались более крепкие мускулы и кости, чтобы справляться с гравитацией без помощи архимедовой силы. Чтобы облегчить переход в это совершенно незнакомое окружение, животные изобрели весьма остроумный трюк. Они забрали с собой, ради юных особей, свое прежнее окружение. По сегодняшний день утроба животного имитирует влажность, текучесть и соленость древнего морского окружения. Более того, концентрация солей в крови и других телесных жидкостях млекопитающих замечательным образом соответствует концентрации солей в океане. Мы вышли из океана более 400 миллионов лет тому назад, но никогда не расставались с морской водой. Мы и теперь обнаруживаем ее в своей крови, поте и слезах. Другое важное нововведение, которое стало существенным для жизни на суше, касалось регуляции содержания кальция. Кальций играет центральную роль в метаболизме всех ядерных клеток. В частности, он необходим для функционирования мышц. Для того чтобы эти метаболические процессы работали, количество кальция должно очень точно поддерживаться на определенных уровнях, гораздо более низких, чем в морской воде. Поэтому морским животным с самого начала пришлось непрерывно удалять весь избыточный кальций. Ранние животные просто выделяли свои кальциевые отходы, иногда нагромождая из них массивные коралловые рифы. По мере того как эволюционировали более крупные животные, они стали накапливать кальций вокруг и внутри себя, и эти отложения в конце концов превратились в раковины и скелеты. Подобно тому как сине-зеленые бактерии преобразовали токсичный загрязнитель, кислород, в жизненно важный ингредиент своей дальнейшей эволюции, так ранние животные преобразовали другой серьезный загрязнитель, кальций, в строительный материал для новых структур, которые давали им огромные преимущества в ходе отбора. Раковины и другие твердые части использовались для защиты от хищников, тогда как скелеты, впервые появившиеся у рыб, впоследствии эволюционировали в важные поддерживающие структуры всех крупных животных. Начало так называемого кембрийского периода (около 580 млн. лет назад) отмечено таким изобилием ископаемых пород с красивыми и четкими отпечатками раковин, твердых покровов и скелетов, что палеонтологи долгое время считали эти кембрийские породы свидетельствами начала жизни. Иногда их даже рассматривали как божественные следы первых актов творения. Лишь в последние три десятилетия следы микрокосма стали обнаруживать в так называемых «химических ископаемых»45. Эти находки убедительно показывают, что зарождение жизни опережает кембрийский период почти на три миллиарда лет. Эволюционные эксперименты с отложениями кальция привели к огромному разнообразию форм — трубчатые «морские спринцовки» со спинным хребтом, но без костей; рыбообразные создания с внешним панцирем, но без челюстей; рыбы, дышащие как в воде, так и в атмосфере, и многие другие. Первые позвоночные со спинным хребтом и черепным костным скелетом, защищающим нервную систему, вероятно, появились около 500 миллионов лет назад. Среди них были предки рыб с легкими короткими плавниками, с челюстями и головой как у лягушки; они ползали вдоль берега и в конце концов эволюционировали в первых амфибий. Амфибии (земноводные) — лягушки, жабы, саламандры и тритоны — служат эволюционным связующим звеном между водными и сухопутными животными. Это первые наземные позвоночные, но даже сегодня они начинают свой жизненный цикл как головастики, дышащие в воде. Первые насекомые вышли на берег примерно в то же время, что и амфибии, и, возможно, даже побудили некоторых рыб последовать за собой, представляя для них лакомую пищу. На суше насекомые породили неимоверное разнообразие видов. Малые размеры и высокая скорость размножения позволяли им приспосабливаться почти к любой окружающей среде, развивая фантастическое разнообразие телесных структур и режимов жизни. Сегодня известно около 750 000 видов насекомых, в три раза больше, чем всех остальных видов животных вместе взятых. В течение 150 миллионов лет после выхода из моря амфибии эволюционировали в рептилий, обладавших значительными преимуществами при отборе — мощными челюстями, кожей, защищающей от засухи, и, что важнее всего, новым типом откладываемых яиц. Как это станут позже делать млекопитающие в своей утробе, рептилии заключили прежнюю среду обитания в большие яйца, внутри которых их отпрыски могли полностью подготовиться к жизненному циклу на суше. Вооруженные этими инновациями, рептилии быстро завоевали сушу и образовали множество разновидностей. Многие виды ящериц, которые существуют до сих пор, являются потомками этих древних рептилий. Эволюция растений и животных Млн лет назад Стадии эволюции 700 ранние животные 620 зачатки мозга у животных 580 раковины и скелеты 500 позвоночные 450 растения выходят на сушу 400 амфибии и насекомые выходят на сушу 350 семенные папоротники 300 грибы 250 рептилии 225 хвойные и динозавры 200 млекопитающие 150 птицы 125 цветковые растения 70 вымирание динозавров 65 ранние приматы 35 мартышки 20 обезьяны 10 человекообразные обезьяны 4 прямоходящие «южные обезьяны» Пока первое поколение рыб выбиралось из воды и превращалось в амфибий, на суше уже процветали кустарники и деревья, и когда амфибии превратились в рептилий, они очутились в густых тропических лесах. В это же время вышел на сушу третий тип многоклеточных организмов — грибы. Грибы похожи на растения, но в то же время столь отличны от них, что были выделены в особое царство, проявляющее ряд замечательных свойств46. У них отсутствует зеленый хлорофилл для фотосинтеза, они не едят и не переваривают, но поглощают нужные питательные вещества непосредственно в форме химических соединений. В отличие от растений, грибы не обладают сосудистой системой для формирования корней, стеблей и листьев. У них есть вполне различимые клетки, которые могут содержать несколько ядер и отделяются друг от друга тонкими стенками, сквозь которые свободно протекает клеточная жидкость. Грибы появились более 300 миллионов лет назад и распространялись через тесную совместную эволюцию с растениями. Фактически все растения, произрастающие на Земле, опираются на помощь крошечных грибков, которые живут в их корнях и обеспечивают поглощение азота. В лесу корни всех деревьев взаимосвязаны через обширную грибковую сеть, которая временами прорывается на поверхность в виде лесных грибов. Без грибов не могли бы существовать первобытные тропические леса. Через тридцать миллионов лет после появления первых рептилий одна из их родовых ветвей эволюционировала в динозавров (греческий термин, в переводе означающий «ужасные ящерицы»), бесконечное очарование которых, похоже, признают люди всех возрастов. Динозавры отличались огромным разнообразием размеров и форм. Некоторые из них обладали панцирем, закрывавшим тело, и костяными наростами — как современные черепахи или носороги. Одни были травоядными, другие — плотоядными. Подобно другим рептилиям, динозавры откладывали яйца. Многие из них строили гнезда, а некоторые даже развили крылья и в итоге, около 150 миллионов лет назад, эволюционировали в птиц. Во времена динозавров распространение рептилий шло полным ходом. Суша и воды были заселены змеями, ящерицами и морскими черепахами, а также морскими змеями и несколькими видами динозавров. Примерно 70 миллионов лет назад динозавры и множество других видов внезапно исчезли, вероятнее всего, в результате падения на Землю огромного метеорита около 7 миль в поперечнике. Катастрофический взрыв вызвал огромное облако пыли, которое на длительный период затмило солнечный свет и привело к критическому изменению погодных паттернов на всей Земле; этих перемен огромные динозавры не смогли пережить. Забота о молодом поколении Около 200 миллионов лет назад из рептилий эволюционировали теплокровные позвоночные; они образовали новый класс животных, из которого в конце концов выделились наши предки, приматы. Женские особи этих теплокровных животных уже не заключали зародышей в яйца, а вынашивали их внутри своего тела. После рождения молодь была относительно беспомощной, и матерям приходилось опекать своих детенышей. Поскольку этот класс животных отличался особым поведением, включающим вскармливание молоком из молочных желез, он получил название млекопитающие. Примерно 50 миллионов лет спустя другие потомки теплокровных позвоночных, птицы, тоже начали вскармливать и обучать своих беззащитных отпрысков. Первые млекопитающие были маленькими ночными животными. Тогда как рептилии, неспособные регулировать температуру тела, были инертны в течение холодной ночи, млекопитающие развили в себе способность поддерживать тепло тела на сравнительно постоянном уровне, независимо от окружающих условий. Благодаря этому они и ночью сохраняли внимательность и активность. К тому же они преобразовали часть своих кожных клеток в шерсть, что еще больше выделило их среди других видов и позволило мигрировать из тропиков в более холодные области. Ранние приматы, известные как прозимианы («предмартышки»), эволюционировали в тропиках около 65 миллионов лет назад из насекомоядных млекопитающих, которые обитали на деревьях и напоминали белок. Сегодняшние прозимианы — это маленькие лесные животные, большей частью ночные; они по-прежнему живут на деревьях. Чтобы ночью перепрыгивать с ветки на ветку, эти ранние обитатели деревьев выработали острое зрение, и у некоторых видов глаза постепенно смещались к плоскости лба, что было решающим для освоения трехмерного видения — важнейшее преимущество для оценки расстояния между деревьями. Другие хорошо известные особенности приматов, связанные с мастерством лазания но деревьям, — цепкие руки и ноги, далеко отстоящий большой палец руки и большие пальцы ног. В отличие от других животных, прозимианы не были анатомически специализированы и поэтому им постоянно угрожали враги. Тем не менее они компенсировали отсутствие специализации значительным развитием ловкости и интеллекта. Угроза со стороны врагов, постоянная жизнь в бегах и ночная активность побуждали их к сотрудничеству и привели к формированию социального поведения, которое характерно для всех высших приматов. Кроме того, привычка охранять себя, издавая частые пронзительные крики, постепенно развилась в общение посредством голоса. Большинство приматов — насекомоядные или вегетарианцы, питающиеся орехами, плодами и травами. Временами, когда на деревьях недоставало орехов и плодов, ранним приматам приходилось покидать спасительные ветви и спускаться на землю. Напряженно высматривая врагов поверх высокой травы, они на короткие промежутки времени принимали вертикальную позицию, чтобы потом опять вернуться в согнутое положение; так ведут себя бабуины и по сей день. Способность стоять прямо — пусть и недолго — оказалась важнейшим преимуществом в ходе естественного отбора, поскольку позволяла использовать руки для собирания пищи, держать палку или бросать камни, защищая себя. Со временем их ступни стали более плоскими, ловкость рук повысилась, использование примитивных орудий труда и защиты стимулировало развитие мозга, и, таким образом, некоторые из прозимианов эволюционировали в мартышек и обезьян. Эволюционная линия мартышек ответвилась от линии прозимианов около 35 миллионов лет назад. Мартышки — это дневные животные, как правило отличающиеся от прозимианов более плоскими и выразительными лицами и обычно передвигающиеся на четырех конечностях. Около 20 миллионов лет назад от линии мартышек отделилась линия обезьян, а еще через десять миллионов лет в свои права вступили наши непосредственные предки, человекообразные обезьяны — орангутанги, гориллы и шимпанзе. Все обезьяны — лесные обитатели, и большинство из них проводят время, по крайней мере частично, на деревьях. Гориллы и шимпанзе, наиболее «приземленные» из всех обезьян, путешествуют на своих четырех, используя ходьбу на кулаках, т. е. опираясь на суставы пальцев передних конечностей. Большинство обезьян способны преодолевать небольшие дистанции и на двух ногах. Подобно людям, обезьяны обладают Широкой плоской грудной клеткой и конечностями, приспособленными к широкому пространственному диапазону действий. Это позволяет им передвигаться по деревьям, попеременно захватывая ветки руками, на что мартышки не способны. Мозг человекообразных обезьян устроен гораздо сложнее, чем у мартышек, и, следовательно, они обладают более высоким интеллектом, чем последние. Для человекообразных обезьян характерна способность использовать и, в ограниченной степени, даже изготавливать орудия труда. Около четырех миллионов лет назад некий вид шимпанзе в африканских тропиках эволюционировал в вертикально передвигающуюся обезьяну. Этот вид приматов, вымерший миллион лет спустя, был очень похож на других человекообразных, но, благодаря прямой походке, его классифицировали как «гоминида», что, согласно Линн Маргулис, совершенно необоснованно с чисто биологической точки зрения: Объективные ученые, если бы они были китами или дельфинами, поместили бы людей, шимпанзе и орангутангов в одну таксономическую группу. Не существует физиологических оснований для выделения человеческих существ в отдельный род… Человеческие существа и шимпанзе имеют гораздо больше общего, чем два произвольно выбранных рода жуков. Несмотря на это, животные с прямой походкой и свободно свисающими руками незаслуженно классифицируются как гоминиды… а не обезьяны47. Приключения человека Следуя за раскрытием жизни на Земле от самых его начал, мы не можем не испытывать особого волнения, когда подходим к той стадии, на которой первые приматы встают на две ноги, — хотя, вероятно, это волнение научно необоснованно. По мере того как мы узнаем, как рептилии эволюционировали в теплокровных позвоночных, которые заботятся о своем потомстве; как первые приматы развивали плоские ногти, далеко отстоящие большие пальцы и голосовое общение; и как обезьяны развивали «человеческую» грудную клетку и руки, сложный мозг и способность изготавливать орудия труда, — мы можем проследить постепенное возникновение человеческих особенностей. А подойдя к стадии прямоходящих обезьян, освободивших свои руки, мы чувствуем, что здесь по-настоящему начинается эволюционное приключение человека. Чтобы подробно его исследовать, мы должны снова изменить свой временной масштаб — на этот раз с миллионов лет на тысячи. Прямоходящие обезьяны, которые вымерли около 1,4 миллиона лет назад, принадлежали роду австралопитеков. Это название, состоящее из латинского australis(«южный») и греческого pithekos(«обезьяна»), означает просто «южная обезьяна». Вид был так назван в честь первого обнаружения в Южной Африке ископаемых останков особи этого вида. Старейшие ископаемые образцы этих южных обезьян известны как Australopithecusafarensis, по имени области Афар в Эфиопии, где они были найдены. Там же был обнаружен и знаменитый скелет, которому дали имя «Люси». Это были приматы легкого телосложения, вероятно 4,5-футового роста и, предположительно, с интеллектом современного шимпанзе. После почти миллиона лет генетической устойчивости, примерно три-четыре миллиона лет назад, первые разновидности южной обезьяны эволюционировали в более крепко сложенных существ. Сюда входят две разновидности людей, которые в течение нескольких сотен тысяч лет сосуществовали с южными обезьянами в Африке, пока последние не вымерли. Важное различие между человеческими существами и другими приматами заключается в том, что детство человеческих отпрысков растягивается на более продолжительный период времени и дети людей, соответственно, достигают половой зрелости и статуса взрослого гораздо позднее, чем любая из обезьян. Если молодь других млекопитающих полностью развивается в утробе и покидает ее уже готовой к жизни во внешнем мире, то наши дети при рождении еще не до конца сформированы и совершенно беспомощны. По сравнению с другими животными кажется, что человеческие детеныши появляются на свет раньше времени. Это наблюдение представляет основу для общепринятой гипотезы о том, что преждевременные роды у некоторых обезьян сыграли решающую роль, дав толчок человеческой эволюции48. Благодаря генетическим изменениям, повлиявшим на временной ход развития особей, незрелорожденные обезьяны могли сохранять свои юношеские особенности дольше, чем другие. Брачные пары таких обезьян, по известному механизму неотении («расширение нового»), давали жизнь еще более недоношенным детенышам, которые сохраняли еще больше черт своей юности. Таким путем могло открыться новое эволюционное направление, в конце концов приведшее к появлению почти безволосого вида, взрослые особи которого во многом походили на зародышей обезьян. Согласно этой гипотезе, беспомощность незрелорожденных детенышей сыграла решающую роль в переходе от обезьян к людям. Новорожденные нуждались в поддержке семьи. Такие семьи формировали сообщества, кочующие племена и поселения, которые заложили основу человеческой цивилизации. Женские особи, как правило, выбирали самцов, которые могли бы позаботиться о них в то время, когда сами они вскармливали и защищали детей. Со временем у самок прекратились сезонные периоды течки, и, поскольку теперь они были сексуально доступны в любое время, самцы, заботящиеся о семье, тоже могли изменить свои сексуальные привычки, упорядочив собственные половые связи в пользу новых социальных условий [1]. В то же время, свобода рук, которые могли изготавливать орудия труда и защиты и бросать камни, стимулировала продолжающееся развитие мозга, характерное для человеческой эволюции, и, возможно, даже внесла свой вклад в развитие языка. Как пишут об этом Маргулис и Саган: Способность бросать камни и оглушать или убивать мелкую добычу вывела первобытных людей в новую эволюционную нишу. Мастерство, необходимое для оценки траектории метательного снаряда, поражения цели на расстоянии, сопряжено с увеличением левого полушария мозга. Развитие языковых возможностей (их тоже связывают с левым полушарием…) могло случайно совпасть с увеличением размеров мозга49. Первые человекообразные потомки южных обезьян появились в Восточной Африке около 2 миллионов лет назад. Это были небольшие стройные существа с заметно развитым мозгом, который обусловил их способности к изготовлению орудий, намного превышавшие возможности предков-обезьян. Поэтому эти первые человеческие виды называют Homohabilis («человек умелый»). Примерно 1,6 миллиона лет назад Homohabilisэволюционировал в более сильный и крупный вид, который продолжал совершенствовать свой мозг. Известный как Homoerectus(«человек выпрямившийся»), этот вид просуществовал более миллиона лет и проявил гораздо большую гибкость, по сравнению со своими предками, приспосабливая свои технологии и образ жизни к широкому диапазону окружающих условий. Существуют свидетельства, что эти первобытные люди около 1,4 миллиона лет назад научились добывать и сохранять огонь. Homoerectusстал первым видом, который покинул уютные африканские тропики и мигрировал в Азию, Индонезию и Европу, укоренившись в Азии около миллиона, а в Европе — около 400 000 лет назад. Вдали от африканской родины первобытным людям пришлось приспосабливаться к исключительно суровым климатическим условиям, которые в итоге оказали значительное влияние на их дальнейшую эволюцию. Вся эволюционная история человеческого рода, от возникновения Homohabilisдо революции в земледелии почти два миллиона лет спустя, совпадает со знаменитыми ледниковыми периодами. В самые холодные эпохи ледяные пласты покрывали обширные области Европы и обеих Америк, а также небольшие площади в Азии. Экстремальные обледенения неоднократно прерывались, и ледники отступали, давая установиться относительно мягкому климату. Вместе с тем колоссальные наводнения, вызванные таянием ледяных масс в межледниковые периоды, представляли серьезнейшую угрозу как для животных, так и для людей. Многие виды животных тропического происхождения вымерли, и их сменили более крепкие, покрытые густой шерстью виды — буйволы, мамонты, бизоны и им подобные, — способные противостоять суровым условиям ледниковых периодов. Первобытные люди охотились на них, используя каменные топоры и дротики, жарили их мясо на кострах, разведенных в пещерах, и использовали шкуры, чтобы защитить себя от жестоких холодов. Охотясь сообща, люди делили и пищу; совместные трапезы стали еще одним катализатором человеческой цивилизации и культуры, породившим и развивавшим с течением времени мифические, духовные и художественные измерения человеческого сознания. Около 400 000 лет назад Homoerectusначал эволюционировать в Homosapiens(«человек разумный») — вид, к которому принадлежат современные люди. Эволюция происходила постепенно; появлялись промежуточные виды, которые принято называть архаическими по отношению к Homosapiens. Примерно 250 000 лет назад Homoerectusвымер; переход к Homosapiensзавершился около 100 000 лет назад в Африке и Азии и примерно 35 000 лет назад — в Европе. Начиная с того времени и до наших дней Homosapiensостается единственным выжившим видом человека. В тот период, когда Homoerectusпостепенно эволюционировал в Homosapiens, в Европе ответвилась еще одна линия, которая примерно 125 000 лет назад развилась в классическую неандертальскую форму. Эволюция человека Лет назадСтадии эволюции 4 млн. Australopithecus afarensis 3,2 млн «Люси» [Australopithecus afarensis) 2.5млннесколькоразновидностейAustralopithecus 2 млнHomo habilis 1,6 млнHomoerectus 1,4 млнАвстралопитеки вымирают 1 млнHomoerectusобосновывается в Азии 400 000Homoerectusобосновывается в Европе; начинает формироваться Homosapiens 250 000архаические формы Homosapiens; Homoerectusвымирает 125 000Homoneanderthalensis 100 000в Африке и Азии окончательно формируется Homosapiens 40 000в Европе окончательно формируется Homosapiens(кроманьонец) 35 000неандертальцы вымирают; Homosapiensостается единственной выжившей разновидностью человека Названный в честь долины Неандер в Германии, где впервые были найдены его останки, этот ярко выраженный вид просуществовал около 90 тысяч лет. Уникальные анатомические черты неандертальцев — они были приземисты и крепки, отличались плотной костью, низко нависающим лбом, тяжелыми челюстями и длинными выступающими вперед зубами — объяснялись, возможно, тем фактом, что они оказались первыми людьми, которые прожили целую эпоху в исключительно холодных условиях, поскольку появились в начале последнего ледникового периода. Неандертальцы закрепились на юге Европы и в Азии, где они оставили следы ритуальных захоронений в пещерах; стены этих пещер украшены разнообразными символами, связанными с культом животных, на которых они охотились. Примерно 35 000 лет назад неандертальцы либо вымерли, либо смешались с формирующимся видом современного человека. Эволюционные приключения человека — это самая новая фаза в раскрытии жизни на Земле и для нас, естественно, наиболее важная и волнующая. Однако с точки зрения Гайи — всей живой планеты, — эволюция человеческих существ является пока лишь коротким эпизодом, который в ближайшем будущем может быстро закончиться. Чтобы продемонстрировать продолжительность пребывания человеческого рода на этой планете, калифорнийский специалист по окружающей среде Дэвид Брауэр придумал весьма остроумную историю, сжав время существования Земли до шести дней библейского творения50. По сценарию Брауэра, Земля была создана в воскресенье в полночь. Жизнь в форме первых бактериальных клеток появляется во вторник утром, примерно в 8:00. Первые два с половиной дня эволюционирует микрокосм; к четвергу, в полночь, он полностью упрочивается, регулируя всю планетарную систему. В пятницу, примерно в 16:00, микроорганизмы изобретают половое размножение, а в субботу, последний день творения, начинают развиваться все крупные формы жизни. Около 1:30 в субботу формируются первые морские животные, за ними два часа спустя следуют амфибии и насекомые. Без пяти пять пополудни появляются огромные рептилии, которые бродят по густым тропикам Земли в течение примерно пяти часов, а затем, приблизительно в 21:45, внезапно вымирают. Тем временем к вечеру, около 17:30, на Земле появляются млекопитающие, а немного погодя, примерно в 19:15, — птицы. Незадолго до 22:00 часть обитающих на деревьях тропических млекопитающих эволюционирует в первых приматов, а через час некоторые из них эволюционируют в мартышек. Около 23:40 появляются человекообразные обезьяны. За восемь минут до полуночи первые южные обезьяны распрямляются и начинают ходить на двух ногах. Пять минут спустя они исчезают. Первый вид людей, Homohabilis, появляется без четырех двенадцать, эволюционирует в Homoerectusчерез полминуты, а в архаические формы Homosapiens— за тридцать секунд до полуночи. Неандертальцы господствуют в Европе и Азии 9 секунд и исчезают за 4 секунды до полуночи. И, наконец, за 11 секунд до полуночи в Африке и Азии и за 5 секунд — в Европе появляются первые современные люди. Вся зафиксированная история человечества длится уже примерно 0,7 секунды. Примерно 35 000 лет назад современный вид Homosapiensпришел на смену неандертальцам Европы и эволюционировал в подвид, известный как кроманьонцы — по названию пещеры на юге Франции, — к которому принадлежат все современные люди. По анатомическим признакам кроманьонцы идентичны нам. Они в полной мере развили язык и произвели настоящую революцию в технологии и художественном творчестве. Тщательно изготовленные орудия из камня и кости, украшения из раковин и слоновой кости и великолепные рисунки на стенах влажных, труднодоступных пещер — яркие свидетельства культурной утонченности этих ранних представителей современной человеческой расы. До недавнего времени археологи полагали, что кроманьонцы совершенствовали свое пещерное искусство постепенно, от грубых и неуклюжих рисунков до блестящих росписей в Ласко, выполненных около 16 000 лет назад. Однако сенсационная находка, обнаруженная в пещере Шове в декабре 1994 года, заставила ученых радикально пересмотреть свои идеи. Эта огромная пещера в провинции Ардеш на юге Франции представляет собой лабиринт подземных помещений, на стенах которых размещено более 300 великолепно выполненных росписей. Стиль напоминает рисунки в Ласко, однако тщательное радиоуглеродное датирование показало, что росписям в Шове не меньше 30 000 лет51. Рисунки, выполненные охрой, углем различных оттенков и красным железняком, представляют символические и мифологические образы львов, мамонтов и других опасных животных; многие из них запечатлены в стремительном беге или прыжке на всю ширину каменных плит пещеры. Специалисты по искусству каменного века были поражены утонченной техникой (тени, динамика фигур, точные углы и др.), которой пользовались пещерные художники для передачи движения и перспективы. Помимо росписей, в пещере находилось множество каменных орудий и ритуальных объектов, в том числе напоминающая алтарь каменная плита, увенчанная черепом медведя. Возможно, самой интригующей находкой оказалось угольное изображение шаманического существа — наполовину человека, наполовину бизона — в самой дальней и темной части пещеры. Неожиданно древняя дата этих великолепных росписей означает, что высокое искусство было неотъемлемой частью эволюции современного человечества с самых первых его шагов. Как отмечают Маргулис и Саган: Одни только эти росписи ясно говорят о присутствии Homosapiensна Земле. Только люди рисуют, только люди придумывают церемониальные походы в отдаленные уголки влажных, темных пещер. Только люди торжественно хоронят своих мертвецов. Поиск исторического предка человека — это поиск сказочника и художника52. Это означает, что верное понимание человеческой эволюции невозможно без понимания эволюции языка, искусства и культуры. Другими словами, сейчас мы должны перенести наше внимание на разум и сознание — третье концептуальное измерение системного взгляда на жизнь. Примечания к главе 10 1. См. Сарга(1982), рр. 116ff. 2. Цитируется там же, р. 114. 3. Margulis(1995). См. ниже, с. 247 и далее. См. выше, с. 223–224. Cm.GouM(1994). Kauffman(1993), pp. 173, 408, 644. Ранние попытки синтеза некоторых из этих элементов см. в Jantsch(1980) и Laszlo(1987). 9. Lovelock(1991),p. 99. 10. См. Margulis and Sagan(1986), pp. 15ff. 11. См. Сарга(1982), рр. 118— 9. См. Margulis and Sagan(1986), p. 75. Там же, р. 16. Там же, р. 89. См. там же. См. там же. Margulis(1995). См. выше, с. 181. Margulis and Sagan(1986), p. 17 Там же, р. 15. Margulis and Sagan(1986); см. также Margulis and Sagan(1995) и Calder(1983). Margulis and Sagan(1986), p. 51. См. выше, с. 111–112; а также Kauffman(1993), pp. 287ff. См. выше, с. 227. Margulis and Sagan(1986), p. 64. См. выше, с. 181. Margulis and Sagan(1986), p. 78. Cm. Lovelock(1991), pp. 80ff. Cm. Margulis(1993), pp. 160ff. См. выше, ее. 183–185. Margulis and Sagan(1986), p. 93. Там же, р. 191. Там же, p. 103. Там же, р. 109. CM.Lovelock(1991), pp. 113ff. См. выше, с. 179 и далее. См. выше, с. 250 и далее. Margulis and Sagan(1986), p. 119. См. выше, с. 183–184. См. Margulis and Sagan(1986), p. 133. См. Thomas(1975), pp. 141ff. Margulis and Sagan(1986), pp. 155ff. Cm. Margulis, Schwartz, and Dolan(1994). Margulis and Sagan(1986), p. 174. Там же, p. 73. Margulis and Sagan(1995), pp. 140ff. Margulis and Sagan(1986), p. 214. См. там же, pp. 208ff. Там же, p. 210. Brower(1995),p. 18. Cm. «New York Times», June 8,1995; Chauvet et al. (1995). Margulis and Sagan(1986), pp. 223–224. Глава 11 Сотворение мира В контексте зарождающейся теории живых систем разум — не вещь, а процесс. Это познание, процесс обучения, тождественный процессу самой жизни. В этом состоит суть теории познания Сантьяго, предложенной Умберто Матураной и Франциско Варелой. Идентификация разума, или обучения, с процессом жизни представляет радикально новую идею в науке, но в то же время является одной из самых глубоких и архаичных интуитивных догадок человечества. В древние времена рациональный человеческий рассудок, или разум, рассматривался просто как один из аспектов нематериальной души или духа. Основное различение проводилось не между телом и разумом, но между телом и душой, или телом и духом. Хотя различие между душой и духом менялось со временем, оба эти термина с самого начала объединяли в себе два понятия — жизненную силу и деятельность сознания2. В древнейших языках обе идеи выражались через метафору дыхания жизни. Действительно, этимологические корни слов «душа» и «дух» во многих языках восходят к «дыханию». Словом, обозначающим «душу» на санскрите (атман), греческом (пневма) и латинском (анима), служит «дыхание». То же относится и к «духу». На латинском {спиритус), греческом {психе) и древнееврейском (руах) это тоже — «дыхание». Всеобщее древнее интуитивное представление, стоящее за этими словами, связывало душу или дух с дыханием жизни. Подобным же образом, концепция познания в теории Сантьяго далеко не ограничивается рациональным разумом, но включает весь процесс жизни. Дыхание жизни — исключительно точная метафора для его описания. Когнитивная наука Подобно концепции «ментального процесса», независимо сформулированной Грегори Бэйтсоном3, теория Сантьяго уходит корнями в кибернетику. Она была разработана в рамках интеллектуального движения, которое подходит к изучению разума и познания с системной, междисциплинарной точки зрения, лежащей за пределами традиционной сферы психологии и эпистемологии. Этот новый подход, который еще не выкристаллизовался в зрелую научную дисциплину, все чаще называют когнитивной наукой4. Кибернетика дала когнитивной науке первую модель обучения. Она исходила из того, что человеческий интеллект подобен компьютерному до такой степени, что познание можно определить как обработку информации, т. е. манипуляцию символами, основанную на наборе правил5. Согласно этой модели, процесс познания включает в себя ментальное представление. Разум здесь осмысливается как компьютер, манипулирующий символами, которые представляют определенные черты мира6. Эта компьютерная модель ментальной деятельности была настолько убедительной и мощной, что господствовала во всех исследованиях когнитивной науки на протяжении более чем тридцати лет. Начиная с 40-х годов вся нейробиология формировалась под воздействием идеи, представляющей мозг в виде устройства для обработки информации. Например, когда исследования зрительной области коры мозга показали, что определенные нейроны реагируют на определенные особенности воспринимаемых объектов — скорость, цвет, контраст и т. д., — сразу возникло представление о том, что эти специализированные нейроны считывают зрительную информацию с сетчатки и передают ее в другие области мозга для дальнейшей обработки. Однако последующие исследования на животных показали, что связывать нейроны с соответствующими характеристиками объектов можно только тогда, когда животное находится под глубоким наркозом и осуществляется строгий контроль над внутренней и внешней средой. Когда животное наблюдают в бодрствующем состоянии и в более привычных для него внешних условиях, его нейронные реакции оказываются более чувствительными ко всему контексту визуального возбудителя и уже не могут быть истолкованы в терминах последовательной обработки информации7. В 70-е годы, когда появилась концепция самоорганизации, компьютерная модель обучения была наконец подвергнута серьезному сомнению. Необходимость свежего критического взгляда на эту доминирующую гипотезу была обусловлена двумя хорошо известными недостатками компьютерного «видения». Первый: обработка информации основана на ряде последовательных правил, применяемых по очереди; | второй: эта обработка локализована таким образом, что повреждение [любой части системы приводит к серьезным нарушениям ее работы в | целом. Обе эти особенности входят в поразительное противоречие с биологическими наблюдениями. Самые обычные визуальные задачи даже крохотными насекомыми решаются быстрее, чем это физически возможно при последовательной обработке; а способность поврежденного мозга к восстановлению и сохранению функционирования в целом хорошо известна всем. Эти наблюдения побуждали к сдвигу фокуса — от символов к связности, от локальных правил к глобальной согласованности, от обычной обработки информации к неожиданным возможностям нейронных сетей. С учетом современного развития нелинейной математики и моделей самоорганизующихся систем, такое смещение внимания обещало новые и интеллектуально волнующие направления исследований. Действительно, в начале 80-х годов модели «связных» нейронных сетей приобрели большую популярность8. Эти модели тесно взаимосвязанных элементов предназначены для одновременного выполнения миллионов операций и проявляют интересные глобальные — внезапно возникающие — свойства. Как поясняет Франциско Варела, «Мозг — это… высоко согласованная система: плотные взаимодействия между его компонентами приводят к тому, что в конечном счете все, что происходит, оказывается функцией того, что делают все компоненты… В результате вся система приобретает внутреннюю согласованность в своих паттернах, хотя мы не можем точно сказать, как это происходит»9. Теория Сантьяго Теория Сантьяго, трактующая познание, возникла в ходе изучения нейронных сетей и с самого начала была связана с концепцией автопоэза, предложенной Матураной10. Познание, или обучение, согласно Матуране, представляет собой деятельность, являющуюся составной частью самосозидания и самоподдержания автопоэзных сетей. Другими словами, обучение — это сам процесс жизни. «Живые системы — это когнитивные системы, — пишет Матурана, — а жизнь — это процесс обучения»11. Обращаясь к нашим трем критериям живых систем — структуре, паттерну и процессу, — мы можем сказать, что жизненный процесс состоит из всех видов деятельности, направленной на непрерывное воплощение системного (автопоэзного) паттерна организации в физической (диссипативной) структуре. Поскольку познание традиционно понимается как процесс получения знания, мы должны описать его как взаимодействие организма с окружающей средой. Именно это и делает теория Сантьяго. Специфическим феноменом, лежащим в основе процесса познания, является структурное сопряжение. Как мы видели, автопоэзная система претерпевает непрерывные структурные изменения, сохраняя в то же время свой паутинообразный паттерн организации. Она сопряжена со своим окружением структурно, т. е. через повторяющиеся взаимодействия, каждое из которых является толчком для структурных изменений в системе12. Тем не менее живая система вполне автономна. Окружение лишь инициирует структурные изменения; оно не определяет и не направляет их. Далее, живая система не просто определяет структурные изменения; она определяет также, какие именно внешние возмущения инициируют их. В этом ключ к пониманию теории Сантьяго. Структурные изменения в системе — это и есть акты познания. Определяя, какие из возмущений, поступающих от внешней среды, становятся начальными толчками перемен, система, как говорят Матурана и Варела, «творит некий мир». В таком случае, познание — это не представление независимо существующего мира, но скорее непрерывное творение мира в процессе жизнедеятельности. Взаимодействие живой системы с окружающей ее средой имеет познавательный характер, и сам процесс жизнедеятельности есть процесс познания. По словам Матураны и Варелы, «жить значит знать»13. Очевидно, что здесь мы имеем дело с радикальным расширением концепции познания и, следовательно, концепции разума. Согласно этому новому подходу, познание охватывает весь процесс жизни — включая восприятие, эмоции и поведение — и не обязательно нуждается в мозге и нервной системе. Даже бактерии воспринимают определенные характеристики своего окружения. Они ощущают химические различия и, соответственно, плывут в сторону сахара и сторонятся кислоты; они ощущают перегрев и избегают его, они движутся к свету или удаляются от него, а некоторые бактерии способны обнаруживать магнитные поля14. Таким образом, даже бактерия творит мир — мир тепла и холода, магнитных полей и химических градиентов. Во всех этих когнитивных процессах восприятие и действие неразделимы, и, поскольку структурные изменения и связанные с ними действия, которые инициируются в организме, зависят от структуры организма, Франциско Варела описывает познание как «воплощенное действие»15. Фактически познание включает в себя два неразрывно связанных вида деятельности: поддержание (и продолжение) автопоэза и созидание мира. Живая система — это многократно взаимосвязанная сеть, чьи компоненты постоянно изменяются, преобразуются и заменяются другими компонентами. Эта сеть отличается исключительной гибкостью и текучестью, что позволяет системе особым образом реагировать на возмущения, или «стимулы», идущие от окружающей среды. Определенные возмущения запускают специфические структурные изменения, т. е. изменения в структуре связи внутри сети. Это распределительный феномен: вся сеть реагирует на выбранное возмущение, перестраивая свои паттерны связи. Разные организмы изменяются по-разному, и со временем каждый организм в процессе развития формирует свое индивидуальное направление структурных изменений. Поскольку эти структурные изменения являются актами познания, развитие всегда ассоциируется с познанием. Фактически развитие и обучение — две стороны одной медали. Оба они суть проявления структурного сопряжения. Не все физические изменения в организме являются актами познания. Когда кролик съедает часть одуванчика или когда зверь получает ранение, эти структурные изменения не определяются и не направляются организмом; они не связаны с выбором и, следовательно, не являются актами познания. Тем не менее эти вынужденные физические изменения сопровождаются другими структурными изменениями (восприятие, реакция иммунной системы и т. д.), которые оказываются актами познания. С другой стороны, не все возмущения, исходящие из окружающей среды, вызывают структурные изменения. Живые организмы реагируют лишь на малую часть возбудителей, воздействующих на них. Общеизвестно, что нам доступны звуки лишь ограниченного диапазона частот; мы часто не замечаем вещи и события в нашем окружении, которые нас не касаются; известно также, что наше восприятие в значительной мере обусловлено рамками наших представлений и культурным контекстом. Другими словами, существует множество возмущений, не вызывающих структурных изменений, поскольку они «чужды» системе. Таким образом, каждая живая система строит свой характерный мир согласно своей характерной структуре. Как отмечает Варела, «разум и мир переживают совместное становление»16. Однако через обоюдное структурное сопряжение, отдельные живые системы составляют части миров друг друга. Они общаются между собой и координируют свое поведение17. Это — экология миров, взращенная взаимно согласованными актами познания. По теории Сантьяго, познание является неотъемлемой частью взаимодействия живого организма с его окружением. Организм не реагирует на раздражающие факторы окружения через линейную причинно-следственную цепочку, но отвечает структурными изменениями в своей нелинейной организационно закрытой автопоэзной сети. Такой тип ответа дает организму возможность поддерживать свою автопоэзную организацию и, следовательно, продолжать свое существование в окружающей среде. Другими словами, когнитивное взаимодействие со своей средой — это взаимодействие разумное. С точки зрения теории Сантьяго, разумность есть проявление богатства и гибкости способов структурного сопряжения организма. Диапазон тех взаимодействий с окружающей средой, которые может осуществлять живая система, определяет ее когнитивную сферу. Эмоции являются естественной частью этой сферы. Например, когда мы отвечаем на оскорбление вспышкой гнева, весь этот паттерн физиологических процессов — пылающее лицо, учащенное дыхание, дрожь и т. п. — является частью познания. И новейшие исследования убедительно подтверждают, что каждый когнитивный акт эмоционально окрашен18. По мере того как возрастает сложность живого организма, расширяется и его когнитивная сфера. В частности, мозг и нервная система значительно расширяют когнитивную сферу организма, поскольку они значительно увеличивают диапазон и развивают дифференциацию структурных сопряжений. На определенном уровне сложности живой организм структурно сопрягается не только с окружающей средой, но и с самим собой, творя тем самым не только внешний, но и внутренний мир. У человеческих существ созидание внутреннего мира тесно связано с языком, мыслью и сознанием19. Не отображение, не информация Являясь частью единой концепции жизни, разума и сознания, когнитивная теория Сантьяго имеет глубокое значение для биологии, психологии и философии. В частности, что касается ее вклада в эпистемологию — раздел философии, изучающий природу знания о мире, — то это, вероятно, один из самых радикальных и спорных ее аспектов. Уникальная особенность эпистемологии, заложенной в теорию Сантьяго, состоит в том, что она противоречит идее, присущей большинству эпистемологии, но редко выражаемой явным образом, — идее о том, что познание есть отображение, представление независимо существующего мира. Компьютерная модель познания как обработки информации была просто своеобразной формулировкой (основанной на ошибочной аналогии) более общей идеи о том, что мир предопределен и независим от наблюдателя, а познание есть ментальное отображение объективных особенностей этого мира внутри познающей системы. Центральным образом здесь является, согласно Вареле, «познающий агент, заброшенный на парашюте в предопределенный мир» и выделяющий его существенные черты через процесс отображения20. Согласно теории Сантьяго, познание есть не отображение независимого, предопределенного мира, но сотворение нового мира. Конкретным организмом в процессе его жизнедеятельности созидается не мир вообще, а некий конкретный мир, всегда обусловленный структурой организма. Поскольку индивидуальные организмы в рамках одного вида обладают более или менее идентичной структурой, они созидают схожие миры. Мы, люди, кроме этого, сообща пользуемся абстрактным миром языка и мысли и благодаря этому сообща творим наш действительный мир21. Матурана и Варела не считают, что где-то существует некая пустота, из которой мы создаем вещество. Есть материальный мир, но он не обладает никакими предопределенными свойствами. Авторы теории Сантьяго не утверждают, что «ничто не существует»; они утверждают, что «ни одна вещь не существует» независимо от процесса познания. Нет объективно существующих структур; нет заданной территории, карту которой мы могли бы составить: само составление карты порождает особенности территории. Нам известно, например, что кошки или птицы видят деревья совершенно иначе, чем мы, потому что воспринимают свет в другом частотном диапазоне. Таким образом, форма и фактура «деревьев», которые они творят, будут отличаться от наших. Глядя на дерево, мы не изобретаем реальность. Но способы, посредством которых мы устанавливаем форму и размеры объектов и выделяем паттерны из множества получаемых нами сенсорных воздействий, зависят от нашего физического устройства. Как сказали бы Матурана и Варела, способы, которые мы используем для структурного сопряжения с окружающей средой, и, следовательно, мир, который мы творим, зависят от нашей собственной структуры. Вместе с идеей о ментальном представлении независимого мира теория Сантьяго отвергает и идею об информации как некоторой совокупности объективных черт этого независимо существующего мира. По словам Варелы: Мы должны подвергнуть сомнению нашу уверенность в том, что мир предопределен и что познание — это отображение. В контексте когнитивной науки это означает, что мы должны пересмотреть идею о том, что в мире существует готовая информация и ее извлекает познающая система22. Отказ от отображения и информации как основных компонентов процесса познания дается с трудом, поскольку мы привыкли постоянно пользоваться обоими этими понятиями. Символы нашего языка, как разговорного, так и письменного, суть отображения вещей и идей; и в нашей обыденной жизни мы рассматриваем факты (время, даты, сводки погоды, телефон друга) как весьма важные для нас элементы информации. Фактически всю нашу эпоху часто называют «веком информации». Как же, в таком случае, Матурана и Варела могут утверждать, что в процессе познания нет информации? Чтобы понять это на первый взгляд озадачивающее утверждение, мы должны помнить, что у человеческих существ познание включает язык, абстрактное мышление и символические понятия — феномены, другим биологическим видам недоступные. Способность абстрактно мыслить, как мы увидим далее, является ключевой характеристикой человеческого сознания, и, благодаря этой способности, мы действительно можем использовать, и используем, ментальное отображение, символы и информацию. Однако эти элементы процесса познания присущи не всем живым системам. И хотя человек часто использует ментальное отображение и информацию, наш познавательный процесс построен не на них. Для того чтобы правильно оценить эти идеи, нам очень полезно будет более пристально рассмотреть то, что подразумевается под «информацией». Обыденное представление сводится обычно к тому, что информация — это «нечто, находящееся где-то», а мозг собирает и обрабатывает это нечто. Однако таким элементом информации является число, имя или краткое сообщение, которое мы извлекаем из всей сети взаимоотношений, из контекста, в который оно заключено и который сообщает ему смысл. Всякий раз, когда такой «факт» заключен в устойчивом контексте и встречается нам с высокой регулярностью, мы можем абстрагировать его от контекста, поставить его в соответствие со значением, присущим ему внутри этого контекста, и назвать это «информацией». Мы настолько привыкли к таким абстрактным операциям, что склонны верить, что значение содержится в элементе информации, а не в контексте, из которого он был извлечен. Например, в красном цвете нет ничего «информативного», за исключением того, что, будучи включенным в культурную сеть соглашений и технологическую сеть дорожного движения, он ассоциируется с остановкой на перекрестке. Если бы люди из какой-то иной культуры приехали в один из наших городов и увидели красный свет светофора, он, скорее всего, не имел бы для них никакого смысла. Передачи информации не произошло бы. Подобным же образом время дня и дата абстрагируются нами от сложных понятий и идей, связанных с солнечной системой, астрономическими наблюдениями и культурными условностями. Эти соображения применимы и к генетической информации, закодированной в ДНК. Варела поясняет, что понятие генетического кода было абстрагировано от лежащей в его основе метаболической сети, в которой только и имеет смысл этот код: Долгие годы биологи рассматривали протеиновые последовательности как инструкции, закодированные в ДНК. Очевидно, однако, что триплеты ДНК могут заранее определять содержание аминокислот в протеине только в том случае, если они включены в клеточный метаболизм, то есть в тысячи ферментных «правил» в сложной химической сети. И только благодаря возникновению внезапных регулярностей в такой сети в целом, мы можем вынести за скобки этотметаболический фон и тогда уже рассматривать триплеты как коды для аминокислот23. Матурана и Бэйтсон Отказ Матураны от идеи о том, что познание включает ментальное отображение независимого мира, служит ключевым различием между его концепцией процесса познания и теорией Грегори Бэйтсона. Матурана и Бэйтсон, практически одновременно и независимо друг от друга, пришли к революционной идее отождествления процесса познания с процессом жизни24. Но они подошли к ней с совершенно разных сторон: Бэйтсон основывался на своем глубоком интуитивном понимании природы разума и жизни, подкрепленном тщательными наблюдениями над живым миром; Матурану вели его попытки определить — на основе нейробиологических исследований — паттерн организации, присущий всем живым системам. Бэйтсон, работая в одиночку, годами оттачивал свои «критерии ментального процесса», но так и не развил их в теорию живых систем. Матурана, в противоположность ему, сотрудничал с другими учеными в разработке теории организации живого, которая обеспечила теоретическую основу для понимания процесса познания как процесса жизни. Как отмечает в своей обширной статье «Понять Бэйтсона и Матурану» исследователь социальной сферы Пол Делл, Бэйтсон сосредоточился исключительно на эпистемологии (природе знания) в ущерб онтологии (природе бытия): Для Бэйтсона онтология остается «нехоженой дорогой»… У эпистемологии Бейтсона нет онтологии, которая могла бы стать ее основанием… Я убежден, что труды Матураны содержат как раз ту онтологию, которую Бэйтсон так и не разработал25. Изучение бэйтсоновских критериев ментального процесса показывает, что они распространяются как на структуру, так и на паттерны живых систем; возможно, из-за этого многие ученики Бэйтсона находили их достаточно сложными для понимания. Внимательный анализ этих критериев выявляет также заложенное в их основу верование, что познание сводится к ментальному отображению объективных характеристик мира в познающей системе26. Бэйтсон и Матурана, независимо друг от друга, разработали революционную концепцию разума, основанную на кибернетике. Бэйтсон способствовал развитию этой традиции еще в 40-е годы. Возможно, именно его увлечение кибернетическими идеями в период их становления привело к тому, что Бэйтсону так и не удалось выйти за пределы компьютерной модели познания. Матурана же, в отличие от него, отказался от этой модели и разработал теорию, в которой познание рассматривается как акт «сотворения мира», а сознание — как феномен, тесно связанный с языком и абстрактным мышлением. Пересмотр компьютерной модели На предыдущих страницах я неоднократно подчеркивал различия между теорией Сантьяго и компьютерной моделью познания, разработанной в рамках кибернетики. Теперь было бы полезно еще раз взглянуть на компьютеры в свете нашего нового понимания познания, чтобы развеять дымку недоразумений, окутывающую «компьютерный интеллект». Компьютер обрабатывает информацию. Это означает, что он манипулирует символами на основе определенных правил. Символы представляют собой определенные элементы, загружаемые в компьютер извне; в ходе обработки информации изменений в структуре машины не происходит. Физическая структура компьютера неизменна, она определена замыслом разработчика и конструкцией. Нервная система живого организма функционирует существенно иначе. Как мы видели, она взаимодействует со своим окружением, постоянно изменяя свою структуру таким образом, что в каждый определенный момент ее физическая структура является записью предыдущих структурных изменений. Нервная система не обрабатывает информацию из внешнего мира, но, наоборот, творит некий мир в процессе познания. В человеческом познании используется язык и абстрактное мышление и, следовательно, символы и ментальные отображения; но абстрактная мысль — это лишь малая часть человеческого познания, и, вообще говоря, она не служит основой для наших повседневных решений и действий. Человеческие решения никогда не бывают в полной мере рациональными, зато всегда окрашены эмоциями; человеческая мысль всегда погружена в телесные ощущения и процессы, которые вносят свой вклад в полный спектр познания. В книге «Компьютеры и познание» исследователи компьютеров Терри Уиноград и Фернандо Флорес подчеркивают, что рациональная мысль отфильтровывает и отбрасывает подавляющую часть когнитивного спектра и тем самым вызывает «слепоту абстракции». Подобно шорам, термины, принятые нами для самовыражения, ограничивают диапазон нашего взгляда на мир. В компьютерной программе, как поясняют Уиноград и Флорес, различные цели и задачи формулируются в терминах ограниченного набора объектов, свойств и операций; этот набор и воплощает ту слепоту, которая приходит вместе с абстракциями, необходимыми для создания программ. Однако: Существуют ограниченные типы задач, в которых эта слепота не исключает достаточно разумного поведения. Например, многие игры предполагают прямое применение… таких программ, которые позволяют переигрывать соперника-человека… Это те области, в которых идентификация требуемых характеристик весьма прямолинейна, а природа решений имеет четкий и ясный характер27. Большая путаница вызвана тем, что компьютерщики используют слова «интеллект», «память» и «язык» для описания компьютеров, тем самым как бы уравнивая эти понятия с человеческими феноменами, хорошо известными из повседневного опыта. Это серьезная ошибка. Например, самая суть разума заключается в том, чтобы действовать наилучшим образом в условиях неопределенной проблемы и неочевидных решений. Разумное человеческое поведение в таких ситуациях основано на здравом смысле, накопленном из жизненного опыта. Здравый смысл, однако, недоступен компьютерам из-за слепоты абстракции и неизбежной ограниченности формальных операций; поэтому и невозможно запрограммировать компьютер на разумность28. Одновременно с идеей искусственного интеллекта появился и великий соблазн запрограммировать компьютер на понимание человеческого языка. Однако после нескольких десятилетий тщетной работы над этой проблемой изобретатели АИ (автоматического интерпретатора) начинают понимать, что все их усилия обречены на неудачу: компьютерам не дано в более или менее достаточной степени понять человеческий язык29. Причина в том, что язык вложен в паутину социальных и культурных условностей, которая содержит и негласный контекст смысла. Мы понимаем этот контекст, потому что он эквивалентен нашему здравому смыслу, но компьютер нельзя запрограммировать на здравый смысл и, следовательно, на понимание языка. Это положение может быть проиллюстрировано множеством простых примеров, вроде текста, приведенного Терри Уиноградом: «Томми только что подарили новый набор кубиков. Он как раз открывал коробку, когда вошел Джимми». Как поясняет Уиноград, компьютер ни за что не догадается, что лежит в коробке, мы же сразу предполагаем, что в ней лежат новые кубики Томми. Мы-то знаем, что подарки обычно приносят в коробках и что самое естественное в этом случае — открыть коробку. И, что еще более важно, мы полагаем, что два предложения в тексте взаимосвязаны, тогда как компьютер не видит смысла в том, чтобы связывать коробку с кубиками. Другими словами, наша интерпретация этого простого текста основана на некоторых связанных со здравымсмыслом предположениях и ожиданиях, недоступных компьютеру30. Тот факт, что компьютер не может понять язык, отнюдь не означает, что он не может быть запрограммирован на распознавание простых лингвистических структур и манипуляции с ними. Действительно, в последние годы в этой области был достигнут значительный прогресс. Сегодня компьютер может распознавать несколько сотен слов и фраз, и этот базовый словарь продолжает расширяться. Так, машины все чаще используются для взаимодействия с людьми посредством структур человеческого языка и выполнения ограниченного круга заданий. Например, я могу позвонить в свой банк и запросить информацию о моем текущем счете; компьютер, если он получит также особый кодовый сигнал, сообщит мне состояние баланса, номера и суммы последних выплат и вкладов и т. п. Такое взаимодействие, предполагающее комбинацию простых произнесенных слов с набранным кодовым номером, очень удобно и полезно; но из этого вовсе не следует, что банковский компьютер понимает человеческий язык. К сожалению, налицо поразительный диссонанс между критическими оценками АИ и радужными проектами компьютерной индустрии (последние явно мотивированы коммерческими интересами). Новейшая волна самых восторженных обещаний исходит от так называемого «проекта пятого поколения», запущенного в Японии. Анализ его грандиозных планов показывает, тем не менее, что они не более реальны, чем аналогичные предыдущие проекты, хотя вполне вероятно, что в рамках программы будет создано немало полезных побочных продуктов31. Центральной идеей проекта пятого поколения и других подобных исследовательских программ служит разработка так называемых «экспертных систем», ориентированных на то, чтобы соперничать с экспертами-людьми в решении определенных задач. Здесь мы опять сталкиваемся с неудачным использованием терминологии. Как отмечают Уиноград и Флорес: Называть программу «экспертом» — значит вводить в заблуждение точно так же, как и называть ее «разумной» или говорить, что она «понимает». Такое неадекватное представление может быть полезным для тех, кто пытается обеспечить финансирование своих исследований или продавать подобные программы, но оно может вызвать необоснованные ожидания у тех, кто пытается их использовать32. В середине 80-х философ Хьюберт Дрейфус и исследователь компьютеров Стюарт Дрейфус предприняли тщательное исследование экспертизы, проводимой людьми, и сопоставили ее с компьютерными экспертными системами. Вот что они обнаружили: …следует расстаться с традиционным убеждением, что новичок учится на частных случаях и лишь по мере приобретения профессионального мастерства начинает абстрагировать и усваивать все более тонкие законы… Приобретение мастерства происходит как раз в противоположном направлении — от абстрактных законов к особым случаям. Похоже, что новичок делает умозаключения, используя законы и факты точно так же, как и эвристически запрограммированный компьютер, однако при наличии таланта и с приобретением соответствующего опыта новичок превращается в эксперта, который интуитивно видит, что нужно делать, не пользуясь законами33. Это замечание показывает, почему экспертные системы никогда не достигают уровня экспертов-людей: последние действуют не по жесткой системе правил, а на основе интуитивного восприятия всей совокупности фактов. Дрейфус и Дрейфус отмечают также, что экспертные системы практически проектируются на основе опроса экспертов-людей, владеющих знанием соответствующих правил. Когда это делается, эксперты чаще всего формулируют те законы, которые запомнили со времен ученичества, но перестали использовать, став профессиональными экспертами. Если эти законы ввести в компьютер, результирующая экспертная система будет копировать новичка, но никогда не сможет соперничать с настоящим экспертом. Когнитивная иммунология Вероятно, наиболее важные практические применения теория Сантьяго нашла в нейробиологии и иммунологии. Как уже отмечалось, новый t взгляд на познание существенно проясняет загадку вековой давности о взаимосвязи между разумом и мозгом. Разум представляет собой не вещь, а процесс — процесс познания, тождественный процессу жизни. 1озг является специфической структурой, с помощью которой этот процесс осуществляется. Таким образом, взаимосвязь между разумом и мозгом — это взаимосвязь между процессом и структурой. Мозг никоим образом не является единственной структурой, вовлеченной в процесс познания. Становится все более очевидным, что иммунная система человека, равно как и других позвоночных, представляет собой сеть не менее сложную и переплетенную, чем нервная система, и выполняет не менее важные координирующие функции. Классическая иммунология рассматривает иммунную систему как защитную систему тела, направленную вовне; ее часто описывают с помощью военных метафор — армии белых кровяных клеток, генералов, солдат и т. д. Последние открытия Франциско Варелы и его коллег из Парижского университета бросают серьезный вызов этой концепции34. Сегодня многие исследователи убеждены, что классический подход с его военными метафорами был одним из главных камней преткновения на пути к разгадке автоиммунных заболеваний, таких, как СПИД. В отличие от нервной системы, сосредоточенной и связанной через анатомические структуры, иммунная система рассеяна в лимфатической жидкости, проникающей в каждую отдельную ткань. Ее компоненты — класс клеток, именуемых лимфоцитами и широко известных как белые кровяные клетки, — очень быстро передвигаются и вступают в химические связи друг с другом. Лимфоциты представляют собой группу на редкость разнообразных клеток. Каждый их тип отличается особыми молекулярными маркерами — антителами, которые выступают над поверхностью этих клеток. Человеческое тело содержит миллиарды белых кровяных клеток различного типа, которые обладают чрезвычайной способностью химически связывать любой молекулярный профиль в окружающей их среде. Согласно традиционной иммунологии, лимфоциты обнаруживают вторгшийся агент, антитела прикрепляются к нему и таким образом его нейтрализуют. Такая последовательность означает, что белые кровяныеклетки распознают чужие молекулярные профили. Более детальные исследования показывают, что этот процесс предполагает также некоторую форму обучения и запоминания. В классической иммунологии, однако, такие представления используют чисто метафорически, не связывая их с каким-либо реальным когнитивным процессом. Недавние исследования показали, что в нормальных условиях антитела, циркулирующие во всем теле, прикрепляются ко многим (если не ко всем) типам клеток, включая и самих себя. Вся система скорее напоминает сеть, множество людей, разговаривающих между собой, но не воинов, высматривающих врага. Постепенно иммунологи были вынуждены изменить свое представление, смещаясь от иммунной системы к иммунной сети. Этот сдвиг представлений оказался серьезной проблемой для классической школы. Если иммунная система — это сеть, компоненты которой связываются друг с другом, и если антитела призваны уничтожать то, с чем они связываются, то выходит, что мы должны разрушать сами себя. Очевидно, что мы этого не делаем. Похоже, что иммунная система все-таки способна отличать клетки собственного тела от чуждых агентов, себя от несебя. Но поскольку, по классической теории, распознать чуждый агент для антитела означает химически прикрепиться к нему и тем самым нейтрализовать его, то остается загадкой, каким образом иммунная система может распознавать собственные клетки, не нейтрализуя их, т. е. не разрушая их функционально. Более того, с традиционной точки зрения, иммунная система может развиваться только тогда, когда происходят внешние возмущения, на которые она реагирует. Если нет атак, антитела не развиваются. Последние эксперименты показали, однако, что животные, полностью защищенные от болезнетворных агентов, все же развивают полноценную иммунную систему. С новой точки зрения, это вполне естественно, поскольку основная задача иммунной системы состоит не в том, чтобы реагировать на внешние угрозы, а в том, чтобы обеспечивать собственную устойчивость35. Варела и его коллеги считают, что иммунную систему следует понимать как автономную когнитивную сеть, которая отвечает за «молекулярную идентичность» тела. Взаимодействуя друг с другом и с другими клетками тела, лимфоциты непрерывно регулируют количество клеток и их молекулярные профили. Иммунная система не просто реагирует на чуждые агенты, но обслуживает важную функцию регулирования клеточного и молекулярного репертуара организма. Как поясняют Франциско Варела и иммунолог Антонио Кутиньо: «Тесный союз иммунной системы и тела позволяет телу поддерживать гибкую и пластичную идентичность на протяжении всей его жизни и реагировать на многочисленные внешние возмущения»36. Согласно теории Сантьяго, когнитивная деятельность иммунной системы обусловлена ее структурным сопряжением с окружающей средой. Когда чуждые молекулы проникают в тело, они вызывают возмущения в иммунной сети, запуская структурные изменения. Результирующая реакция состоит не в автоматическом разрушении чуждых молекул, но в регулировании их уровня в контексте других регулирующих механизмов системы. Реакция может быть различной: она зависит от состояния всей системы. Когда иммунологи вводят в организм большие объемы чуждого агента, как это делается в стандартных экспериментах над животными, иммунная система дает массированный защитный ответ, описанный в классической теории. Однако, замечают Варела и Кутиньо, это в высшей степени искусственная лабораторная ситуация. В своей естественной окружающей среде животное не получает больших порций вредоносных веществ. Малые количества, которые действительно проникают в тело, естественным образом включаются в текущую регуляторную деятельность иммунной сети. С пониманием иммунной системы как когнитивной, самоорганизующейся и саморегулирующей сети тайна различения «свой— чужой» легко раскрывается. Иммунная система не делит клетки на свои и чужие (такое разделение ей и не требуется), поскольку и те и другие являются объектами регулирующих процессов. Тем не менее, если вторжение чуждых агентов оказывается настолько массированным, что они не могут быть включены в регулирующую сеть, как, например, в случае инфекции, они приводят в действие специфические механизмы иммунной системы, которые формируют защитную реакцию. Исследования показали, что широко известный иммунный ответ такого рода включает квазиавтоматические механизмы, практически независимые от когнитивной деятельности сети37. По традиции, иммунология имела дело исключительно с такой «рефлекторной» иммунной деятельностью. Ограничиться этими исследованиями все равно, как если бы мы ограничили исследование мозга изучением рефлексов. Защитная иммунная деятельность очень важна, но в свете новых воззрений она являет собой лишь второстепенную функцию когнитивной деятельности иммунной системы, сосредоточенной преимущественно на внутренних процессах и поддерживающей молекулярную идентичность тела. Область когнитивной иммунологии пока еще переживает период становления, и самоорганизующие свойства иммунных сетей поняты далеко не до конца. Тем не менее некоторые ученые, проявляющие активность в этой новой сфере исследований, уже стали задумываться над многообещающими клиническими применениями в области лечения автоиммунных заболеваний38. Вероятно, терапевтические стратегии будущего будут основаны на понимании того, что автоиммунные заболевания отражают нарушения в функционировании иммунной сети. Эти стратегии, возможно, будут опираться на новые технологии, призванные укрепить сеть, восстанавливая ее связность. Такие технологии, однако, потребуют гораздо более глубокого понимания богатой динамики иммунных сетей, прежде чем их можно будет эффективно применять. В будущем открытия когнитивной иммунологии обещают стать исключительно важными для всей сферы здравоохранения и медицины. По мнению Варелы, утонченный психосоматический взгляд на здоровье («разум-тело») не сможет развиваться, пока мы не привыкнем понимать нервную и иммунную системы как две взаимодействующие когнитивные системы, как два «мозга» в непрерывном диалоге39. Психосоматическая сеть Важный недостающий фрагмент картины был найден в середине 80-х нейробиологом Кэндейс Перт и ее коллегами из Национального института душевного здоровья в Мэриленде. Эти исследователи идентифицировали группу молекул, называемых пептидами, в роли молекулярных посланников, обеспечивающих диалог между нервной и иммунной системами. Фактически Перт и ее коллеги обнаружили, что эти посланники связывают три автономные системы — нервную, иммунную и эндокринную — в единую сеть. Согласно традиционному взгляду, это три отдельные системы и выполняют они разные функции. Нервная система, состоящая из мозга и сети нервных клеток, пронизывающей все тело, представляет собой вместилище памяти, мыслей и эмоций. Эндокринная система, состоящая из желез и гормонов, является основной регулирующей системой тела, контролируя и интегрируя разнообразные телесные функции. Иммунная система, состоящая из селезенки, костного мозга, лимфатических узлов и иммунных клеток, циркулирующих по телу, служит защитной системой тела, отвечающей за цельность ткани и контролирующей заживление ран и механизмы восстановления тканей. Соответственно, эти три системы изучаются тремя отдельными дисциплинами — нейробиологией, эндокринологией и иммунологией. Между тем, новые исследования пептидов убедительно показали, что это концептуальное разделение представляет собой не что иное как исторический артефакт, с которым больше нельзя мириться. Согласно Кэндейс Перт, эти три системы следует рассматривать как нераздельные части единой психосоматической сети40. Пептиды — около 60–70 особых макромолекул — вначале изучались в других контекстах, и им давали разные названия — гормоны, нейропередатчики, эндорфины, факторы роста и т. д. Понадобилось много лет, чтобы увидеть в них единое семейство молекулярных посланников. Эти посланники представляют собой короткие цепочки аминокислот, которые прикрепляются к специфическим рецепторам, в изобилии рассеянным по поверхности всех клеток тела. Поддерживая взаимосвязь между иммунными клетками, железами и клетками мозга, пептиды формируют психосоматическую сеть, пронизывающую весь организм. Пептиды оказываются биохимическим проявлением эмоций; они играют очень важную роль в координирующей деятельности иммунной системы; они связывают и объединяют ментальную, эмоциональную и биологическую деятельность. Решающие перемены в наших представлениях начались в 80-е годы, когда было сделано парадоксальное открытие: определенные гормоны, которые, как предполагалось, производятся железами, оказались пептидами; более того, выяснилось, что они также производятся и хранятся в мозге. С другой стороны, ученые обнаружили, что нейропередатчики, именуемые эндорфинами, которые, как считалось, производятся только в мозге, генерируются также иммунными клетками. По мере того как обнаруживалось все больше и больше пептидных рецепторов, становилось очевидным, что практически любой из известных пептидов может производиться и в мозге, и в различных частях тела. И тогда Кэндейс Перт провозглашает: «Я больше не могу проводить четкое разграничение между мозгом и телом»41. В нервной системе пептиды производятся в нервных клетках и затем перемещаются вниз по аксонам (длинным ответвлениям нервных клеток), где и хранятся на дне в виде крошечных шариков до тех пор, пока соответствующие сигналы не освободят их для деятельности. Эти пептиды играют существенную роль в поддержании связи внутри нервной системы. Традиционно считалось, что передача нервных импульсов происходит через промежутки (синапсы) между соседними нервными клетками. Однако оказалось, что этот механизм не столь важен и используется главным образом для сокращения мускулов. Большинство сигналов, поступающих из мозга, передаются через пептиды, генерируемые нервными клетками. Прикрепляясь к рецепторам вдали от «материнских» нервных клеток, эти пептиды функционируют не только в пределах всей нервной системы, но и в других тканях тела. В иммунной системе белые кровяные клетки не только обладают рецепторами для всех пептидов, но и сами производят пептиды. Пептиды управляют миграционными паттернами клеток и всеми их жизненными функциями. Это открытие, как и успехи когнитивной иммунологии, несомненно, должно найти замечательные терапевтические применения. Перт и ее команда недавно открыли новый многообещающий метод лечения СПИДа, названный Пептидом Т42. Ученые выдвинули гипотезу, что СПИД обусловлен нарушением пептидных связей. Установив, что ВИЧ (вирус иммунодефицита человека) проникает в клетки через определенные пептидные рецепторы, нарушая тем самым функции всей сети, Перт и ее коллеги спроектировали защитный пептид, который прикрепляется к этим рецепторам и блокирует воздействие вируса. (Пептиды возникают естественным образом в теле, но они могут быть также спроектированы и синтезированы). Пептид Т имитирует деятельность естественных пептидов, и, следовательно, он совершенно не токсичен, в отличие от других препаратов против СПИДа. В настоящее время это средство проходит клинические испытания. Если оно окажется эффективным, в лечении СПИДа может произойти настоящая революция. Еще один замечательный аспект недавно признанной психосоматической сети: пептиды оказались биохимическим проявлением эмоций. Большинство пептидов, если не все, влияют на поведение и настроение, и сегодня ученые выдвигают гипотезу, что каждый пептид, вероятно, порождает уникальный эмоциональный тон. Вся группа из 60–70 пептидов, возможно, составляет универсальный биохимический язык эмоций. Нейробиологи традиционно связывали эмоции со специфическими областями мозга, в частности с лимбической системой. И это действительно так. Оказывается, что лимбическая система сильно насыщена пептидами. Однако это не единственная часть тела, где сконцентрированы пептидные рецепторы. Например, весь наш кишечник наполнен пептидными рецепторами. Вот почему мы «чувствуем нутром». Мы буквально ощущаем эмоции в своем кишечнике. Если верно, что каждый пептид передает особое эмоциональное состояние, это означает, что все сенсорные ощущения, все мысли — фактически все телесные функции — окрашены эмоционально, поскольку все они связаны с пептидами. Действительно, ученые наблюдали, что узловые точки нервной системы, через которые осуществляется связь сенсорных органов с мозгом, обогащены пептидными рецепторами, которые фильтруют сенсорные восприятия и определяют их приоритет. Другими словами, все наши восприятия и мысли окрашены эмоциями. Все это, конечно, хорошо известно нам из опыта. Из признания такой психосоматической сети вытекает, что нервная система не структурирована иерархически, как это полагалось раньше. Как отмечает Кэндейс Перт, «белые кровяные клетки — это частицы мозга, путешествующие по всему телу»43. В конечном итоге это означает, что познание есть феномен, сфера действия которого охватывает весь организм. Познание осуществляется через сложную химическую — пептидную — сеть, которая объединяет нашу ментальную, эмоциональную и биологическую деятельность. Примечания к главе 11 См. выше, ее. 191–192. См. Windelband(1901), pp. 232-33. См. выше, с. 190 и далее. См. Varela et al.(1991), pp. 4ff. См. выше, с. 83 и далее. См. Varela et al.(1991), pp. 8, 41. Там же, р. 93–94. См. Gluck and Rumelhart(1990). См. Varela et al.(1991), p. 94. См. выше, с. 114. См. там же. См. выше, с. 237–238. Maturana and Varela(1987), p. 174. См. Margulis and Sagan(1995), p. 179. Varela et al.(1991), p. 200. Там же, р. 177. См. ниже, с. 287 и далее. См. ниже, с. 315–316. См. ниже, с. 310–311. Varela etal.(1991), p. 135. См. ниже, с. 310–311. Varela etal.(1991), p. 140. Там же, p. 101. См. выше, с. 189. Dell(1985). См. Приложение, с. 327 и далее. Winograd and Flores(1991),p. 97. См. там же, pp. 93ff. Там же, pp. 107ff. Там же,p. 113. Там же, pp. 133ff. Там же, р. 132. Dreyfus and Dreyfus(1986),p. 108. См. Varela and Coutinho(1991a). Cm. Varela and Coutinho(1991b). Varela and Coutinho(1991a). Там же. См. Varela and Coutinho(1991b). Франциско Варела, частная беседа, апрель 1991. Pert et al.(1985), Pert(1993). Pert(1989). 42. Cm. Pert(1992), Pert(1995). 43.Pert(1989). Глава 12 Знать о своем знании Для того чтобы отождествить познание со всем процессом жизни — включая восприятия, эмоции и поведение — и понимать его как процесс, который не включает ни передачи информации, ни ментального отображения внешнего мира, мы нуждаемся в радикальном расширении рамок науки и философии. Одна из причин, по которой этот взгляд на разум и познание так трудно принять, состоит в том, что он противоречит нашему повседневному опыту и интуиции. Как человеческие существа, мы часто пользуемся понятием информации и постоянно формируем ментальные отображения людей и объектов из нашего окружения. Между тем эти весьма специфические особенности человеческого познания обусловлены нашей способностью абстрактно мыслить — ключевой характеристикой человеческого сознания. Таким образом, для полной картины общего процесса познания в живых системах нам важно понять, каким образом из когнитивного процесса, характерного для всех живых организмов, возникает человеческое сознание с его абстрактной мыслью и концептуальными понятиями. На последующих страницах термин сознание применяется к такому уровню разума, или познания, когда уже возникло самосознание. Осознание окружающей среды, согласно теории Сантьяго, является свойством познания на всех уровнях жизни. Самосознание, насколько нам известно, присуще только высшим животным и в полной мере проявляется только в человеческом разуме. Будучи людьми, мы осознаем и окружающую среду, и самих себя, и свой внутренний мир. Другими словами, мы осознаем, что мы осознаем. Мы не просто знаем, но и знаем о своем знании. Именно эту особенность самосознания я имею в виду, когда использую термин сознание. Язык и общение В теории Сантьяго самосознание рассматривается в тесной связи с языком, а понимание языка достигается через тщательный анализ общения. Такой подход к пониманию сознания был впервые предложен Умберто Матураной1. Общение, согласно Матуране, это не процесс передачи информации, а координация поведения живых организмов посредством их взаимного структурного сопряжения. Эта взаимная координация поведения является ключевой характеристикой общения всех живых организмов, независимо от наличия у них нервной системы, и становится все более утонченной и сложной по мере нарастания сложности нервных систем. Птичье пение — одна из самых прекрасных форм общения в мире животных. Матурана приводит в качестве поразительного примера брачную песню африканских попугаев. Эти птицы часто обитают в густых лесах, где практически исключена возможность зрительного контакта. В условиях этой окружающей среды пары попугаев формируют и координируют свой брачный ритуал, совместно исполняя особую песню. Случайному слушателю кажется, что каждая из птиц исполняет самостоятельную мелодию, однако при ближайшем рассмотрении, эта песня на самом деле оказывается дуэтом, в котором две птицы попеременно подхватывают и развивают мотивы друг друга. Песня каждой пары неповторима, она не передается потомству. В каждом поколении новые пары создают собственные оригинальные мелодии для брачного ритуала. По словам Матураны: В этом случае (в отличие от многих других птиц) вокальная координация поведения в поющей паре является онтогенетическим феноменом [феноменом развития отдельных особей]… Конкретная мелодия каждой пары птиц этого вида уникальна и связана с историей их спаривания2. Этот понятный и красивый пример подтверждает мысль Матураны о том, что общение, в сущности, есть координация поведения. В других случаях мы склонны описывать общение в семантических терминах, т. е. как обмен информацией, в которой заложен некоторый смысл. Однако согласно Матуране, такие семантические описания являются проекциями наблюдателя-человека. В реальности же координация поведения обусловливается не смыслом, а динамикой структурного сопряжения. Поведение животных может характеризоваться врожденными («инстинктивными») или благоприобретенными чертами, и, соответственно, можно различать инстинктивный и благоприобретенный тип общения. Тип общения, развитый в процессе обучения, Матурана называет лингвистическим. И хотя это еще не язык, с языком его роднит характерная особенность, заключающаяся в том, что одна и та же координация поведения может быть осуществлена посредством взаимодействий различного типа. Как и различные языки человеческого общения, различные виды структурного сопряжения, развитые разными путями, могут привести к одинаковой координации поведения. Именно такое лингвистическое поведение лежит, по мнению Матураны, в основе языка. Лингвистическое общение требует наличия достаточно сложной нервной системы, поскольку оно включает очень сложные формы обучения. Например, когда пчелы сообщают друг другу, где расположены определенные цветы, сплетая в танце сложные паттерны, этот танец отчасти основан на инстинктивном поведении, а частично усвоен в процессе обучения. Лингвистические (или благоприобретенные) компоненты этого танца обусловлены контекстом и социальной историей особи. Пчелы из различных роев танцуют, можно сказать, на разных «диалектах». Даже самые изощренные формы лингвистического общения — например, так называемый «язык» пчел — это еще не язык. Согласно Матуране, язык появляется тогда, когда возникает общение по поводу общения. Другими словами, процесс оязычивания [languaging], как его называет Матурана, знаменует собой координацию координации поведения. Матурана любит иллюстрировать это свойство языка на примере гипотетического общения кошки с хозяином3. Представьте себе, что каждое утро моя кошка мяукает и бежит к холодильнику. Я следую за ней, достаю немного молока, наливаю его в миску, и кошка начинает его лакать. Это и есть общение — координация поведения через постоянное взаимодействие, или взаимное структурное сопряжение. Теперь представьте, что в одно прекрасное утро я не реагирую на мяуканье кошки, так как знаю, что молоко кончилось. Если бы кошка могла сообщить мне что-то вроде: «Эй, я уже мяукнула три раза; где мое молоко?», — это был бы язык. Ее ссылка на предшествующее мяуканье составляла бы сообщение по поводу сообщения и, следовательно, по определению Матураны, квалифицировалась бы как язык. Кошки не умеют использовать язык в этом смысле, но, по-видимому, он доступен человекообразным обезьянам. Американские психологи показали, что шимпанзе способны не только выучить множество стандартных символов языка жестов, но и создавать новые выражения, комбинируя различные жесты4. Так, одна из шимпанзе по имени Люси изобрела несколько знаковых комбинаций: «плод-пить» для арбуза, «пища-плакать-сильный» для редьки и «открывать-пить-есть» для холодильника. Однажды Люси очень расстроилась, увидев, что ее человеческие «родители» собираются уходить. Она обратилась к ним и показала жестами «Люси-плакать». Делая это заявление по поводу своего плача, она явно общалась по поводу общения. «Нам кажется, — пишут Матурана и Варела, — что в этот момент Люси начала оязычиваться»5. Хотя некоторые приматы, очевидно, обладают потенциалом для общения на языке жестов, их лингвистическая сфера крайне ограничена и даже не приближается к богатству человеческого языка. В человеческом языке открывается огромное пространство, в котором слова служат жетонами для лингвистической координации действий, а также для создания понятий об объектах. Например, во время пикника мы можем использовать слова для лингвистического различения, координируя свои действия при накрывании пня скатертью и его сервировке. Кроме того, мы можем также опираться на эти лингвистические различия (т. е. проводить различение между различениями), используя слово «стол» и, тем самым, творя объект. В таком случае объекты, по мнению Матураны, являются лингвистическими различениями лингвистических различений, и, поскольку существуют объекты, мы можем создавать абстрактные понятия — такие как, например, высота стола, — проводя различение различений различения и т. д. Пользуясь терминологией Бэйтсона, можно сказать, что иерархия логических типов возникает вместе с человеческим языком6. Оязычивание Более того. Наши лингвистические различения не изолированы, но существуют «в сети структурных сопряжений, которую мы непрерывно сплетаем через [оязычивание]»7. Значение возникает как паттерн взаимоотношений между этими лингвистическими различениями; таким образом, мы существуем на «семантической территории», созданной нашим оязычиванием. И, наконец, возникает самосознание — когда мы используем представление об объекте и связанные с ним абстрактные понятия для описания самих себя. Таким образом, лингвистическая территория человека простирается еще дальше, включая рефлексию и сознание. Уникальность человеческого бытия заключается в нашей способности непрерывно создавать лингвистическую сеть, в которую вплетены и мы сами. Быть человеком — значит существовать в языке. Через язык мы координируем наше поведение, через язык мы вместе творим мир. «Тот мир, который каждый из нас видит, — пишут Матурана и Варела, — не есть определенный мир, но некий мир, который мы созидаем вместе с другими»8. Человеческий мир зиждется на нашем внутреннем мире абстрактной мысли, понятий, символов, ментальных отображений и самосознания. Быть человеком — значит обладать рефлексивным сознанием: «Узнавая, как мы знаем, мы творим себя»9. В ходе беседы наш внутренний мир понятий и идей, наши эмоции и телесные движения вступают в тесную взаимосвязь, формируя сложную хореографию поведенческой координации. Анализ видеозаписей показал, что каждая беседа включает утонченный и, по большей части, бессознательный танец, в котором последовательность речевых паттернов синхронизируется не только с мельчайшими телесными движениями говорящего, но и с соответствующими движениями слушателя. Оба партнера включены в эту точно синхронизированную последовательность ритмических движений, и лингвистическая координация их взаимно обусловленных действий длится до тех пор, пока поддерживается беседа10. Теория Матураны, трактующая сознание, фундаментально отличается от других подобных теорий тем, что основной ее акцент ставится на язык и общение. С точки зрения теории Сантьяго, модные сегодня попытки объяснить человеческое сознание через квантовые эффекты в мозге или через другие нейрофизиологические процессы обречены на провал. Самосознание и раскрытие внутреннего мира понятий и идей не поддаются объяснению не только на языке физики и химии; они не могут быть поняты даже через биологию или психологию отдельного организма. Согласно Матуране, мы можем понять человеческое сознание только через язык и полный социальный контекст, в который он включен. Сама семантика слова — «сознание» (совместное знание) — предполагает, что речь идет, по существу, о феномене социальном. Полезно сравнить понятие творение мира также с древнеиндийским понятием майя. В ранней индуистской мифологии слово майя означало «магическую творческую силу», с помощью которой созидается мир в божественной игре Брахмана11. Мириады воспринимаемых нами форм созидаются божественным актером и магом, а динамической силой, движителем его игры является карма, дословно означающая «деяние». По прошествии нескольких веков смысл слова майя — одного из важнейших понятий в индийской философии — изменился. Вместо творческой силы Брахмана оно стало обозначать психологическое состояние человека под чарами магической игры. Поскольку мы принимаем материальные формы игры за объективную реальность, не ощущая единства Брахмана, лежащего в основе всех этих форм, постольку находимся под влиянием чар майи. Индуизм отрицает существование объективной реальности. Как и в теории Сантьяго, воспринимаемые нами объекты созидаются через действие. Тем не менее здесь процесс созидания мира происходит в космическом масштабе, а не на уровне человеческого познания. Мир, творимый в индуистской мифологии, не является неким миром определенного человеческого сообщества, но представляет собой конкретный мир магической божественной игры, которая околдовывает всех нас. Первичные состояния сознания В последние годы Франциско Варела исследует еще один подход к сознанию, который, как он надеется, может внести дополнительное измерение в теорию Матураны. Его базовая гипотеза состоит в том, что все высшие позвоночные наделены некой формой первичного сознания, которая, не будучи еще саморефлексивной, уже включает в себя опыт «единого ментального пространства», или «ментального состояния». Многочисленные недавние опыты над животными и людьми показали, что это ментальное пространство обладает многими измерениями; другими словами, оно создается множеством различных функций мозга. И все же это — единый, связный опыт. Например, когда аромат духов вызывает приятные или неприятные ощущения, мы испытываем единое, связное ментальное состояние, в которое включены сенсорные ощущения, воспоминания и эмоции. Это переживание неустойчиво и, как мы знаем из опыта, может быть крайне непродолжительным. Ментальные состояния носят преходящий характер — они непрерывно возникают и исчезают. Однако не представляется возможным испытывать их иначе, чем на некотором конечном отрезке времени. Еще одно важное наблюдение заключается в том, что это эмпирическое состояние всегда «воплощено», т. е. включено в некоторое определенное поле ощущений. В самом деле, большинство ментальных состояний сопровождаются доминирующим чувством, которое дает окраску всему переживанию. Варела недавно опубликовал статью, в которой он выдвигает свою базовую гипотезу и предлагает специфический нейронный механизм для формирования первичных состояний сознания у всех высших позвоночных12. Ключевая идея состоит в том, что переходные эмпирические состояния порождаются резонансным феноменом, известным как фазовая синхронизация, при которой различные разделы мозга соединяются таким образом, что все их нейроны возбуждаются синхронно. Посредством такой синхронизации нейронной деятельности образуются временные клеточные сообщества, в которые могут входить обширные, распределенные по всему организму нейронные системы. Согласно гипотезе Варелы, каждое когнитивное переживание основано на подобном клеточном сообществе, в котором различные типы нейронной активности (сопровождающие сенсорные ощущения, эмоции, память, телесные движения и т. п.) объединяются во временный, но согласованный ансамбль осциллирующих нейронов. Тот факт, что нейронные цепи, как правило, осциллируют в ритмичном режиме, хорошо известен нейробиологам, а последние исследования показали, что эти осцилляции не ограничиваются корой головного мозга, но распространяются на различные уровни нервной системы. Многочисленные эксперименты, упомянутые Варелой в подтверждение этой гипотезы, показывают, что когнитивные эмпирические состояния возникают благодаря синхронизации быстрых осцилляции в гамма- и бета-диапазонах; такие осцилляции обычно очень быстро возникают и затухают. Каждой фазовой синхронизации соответствует характерное время затухания — оно и определяет минимальную длительность переживания. Гипотеза Варелы устанавливает нейробиологическую основу для различения сознательного и бессознательного познания, которую нейробиологи искали с тех пор, как Зигмунд Фрейд открыл человеческое бессознательное13. Согласно Вареле, первичное сознательное переживание, характерное для всех высших позвоночных, не сосредоточено в конкретном разделе мозга и не может быть определено в рамках специфических нейронных структур. Это не что иное, как проявление конкретного когнитивного процесса — преходящая, кратковременная синхронизация разнообразных ритмично осциллирующих нейронных цепей. Человеческое состояние Человеческие существа эволюционировали из прямоходящих южных обезьян (род Australopithecus) около двух миллионов лет назад. Переход от обезьян к людям, как мы узнали из прошлых глав, стимулировали две выразительные особенности развития: беспомощность несформированных новорожденных детей, которые нуждались в поддержке семьи и сообщества, и свобода рук, позволявшая изготавливать и использовать орудия; все это способствовало развитию мозга и, вероятно, эволюции языка14. Теория языка и сознания, предложенная Матураной, позволяет установить связь между этими двумя побудительными эволюционными аспектами. Поскольку язык обеспечивает весьма утонченную и эффективную координацию поведения, эволюция языка позволила первобытным людям значительно повысить уровень сотрудничества и развить семьи, сообщества и племена, что давало им огромные эволюционные преимущества. Решающая роль языка в человеческой эволюции состояла не в возможности обмена идеями, но в расширении возможностей сотрудничества. С ростом многообразия и богатства человеческих взаимоотношений, соответственно, раскрывалось и человечество — его язык, искусство, мысль и культура. В это же время совершенствовалась наша способность к абстрактному мышлению, создавался наш внутренний мир понятий, объектов и образов самих себя. Постепенно, по мере того как этот внутренний мир становился более разнообразным и сложным, мы теряли связь с природой и превращались во все более «отдельных» личностей. Так возник конфликт между целостностью и отдельностью, между телом и душой; поэты, философы и мистики всех времен и народов считали этот конфликт сутью человеческого состояния. Человеческое сознание породило не только пещерные рисунки Шове, Бхагавад-гиту, Бранденбургские концерты и теорию относительности, но также и рабство, охоту на ведьм, холокост, бомбардировку Хиросимы. Среди всех биологических видов наш — единственный, в пределах которого особи уничтожают друг друга ради религиозных, рыночных, патриотических и других абстрактных идей. В буддийской философии ярко представлены некоторые важнейшие проявления человеческого состояния и его корней в языке и сознании15. Экзистенциальное человеческое страдание возникает, по мнению буддистов, когда мы цепляемся за постоянные формы и категории, созданные разумом, вместо того чтобы принять непостоянную, преходящую природу всех вещей. Будда учил, что все постоянные формы — вещи, события, люди или идеи — не что иное как майя. Подобно ведическим пророкам и мудрецам, он пользовался этим древним понятием, но он спустил его с космического уровня, который оно занимает в индуизме, и соединил с процессом человеческого познания, тем самым придав ему свежее, почти психотерапевтическое звучание16. В силу собственного невежества (авидъя), мы делим воспринимаемый мир на отдельные объекты, которые кажутся нам прочными и постоянными, но которые на самом деле переменчивы и преходящи. Цепляясь за свои жесткие категории, не понимая текучести жизни, мы обречены переживать страдание за страданием. Буддистское учение о непостоянстве включает концепцию о несуществовании «я» — постоянного субъекта наших меняющихся переживаний. Главная идея заключается в том, что изолированное, индивидуальное «я» — это иллюзия, всего лишь одно из воплощений майи, интеллектуальное понятие, за которым нет никакой реальности. Цепляние за эту идею изолированного «я» приводит к такой же боли и страданию (дукха), как и приверженность к любой другой застывшей мысленной категории. Когнитивная наука пришла к точно такой же позиции17. Согласно теории Сантьяго, мы творим свое «я» точно так же, как творим объекты. Наше «я», или эго, не существует независимо, но является результатом нашего внутреннего структурного сопряжения. Тщательный анализ веры в независимое, постоянное «я», а также ее последствий — «картезианской тревоги» — привел Франциско Варелу и его коллег к следующему заключению: Наш лихорадочный поиск внутренней твердой почвы — это и есть сущность эго, источник постоянной фрустрации… Поиск внутреннего основания составляет только часть более крупного паттерна поиска, в который входит и наше цепляние за внешнее основание — в форме идеи о предопределенном и независимом мире. Другими словами, наш поиск основания, внутреннего или внешнего, является глубоким источником разочарований и тревог18. Итак, это одна из тяжелейших проблем человеческого состояния. Мы — автономные индивиды, сформированные собственной историей структурных изменений. Мы осознаем себя, осознаем свою индивидуальную идентичность — и все же, когда мы ищем независимую самость в рамках мира нашего опыта, все эти поиски заканчиваются неудачей. Происхождение этой дилеммы определяется нашей тенденцией создавать абстракции изолированных объектов, включая изолированное «я», и потом верить в то, что они принадлежат объективной, независимо существующей реальности. Чтобы преодолеть картезианскую тревогу, нам необходимо мыслить системно, сдвигая свой концептуальный фокус от объектов к взаимоотношениям. Только тогда мы сможем понять, что идентичность, индивидуальность и автономия отнюдь не означают изолированность и независимость. Как напоминают нам Линн Маргулис и Дорион Саган, «независимость — это политический, а не научный термин»19. Сила абстрактного мышления побуждает нас обращаться с естественной окружающей средой — паутиной жизни — так, как если бы она состояла из изолированных частей, предназначенных для эксплуатации различными заинтересованными группами. Более того, мы распространили этот «фрагментный» взгляд и на свое человеческое общество, деля его на различные национальные, расовые, религиозные и политические группы. Вера в то, что все эти фрагменты — внутри нас самих, в нашей окружающей среде и в нашем обществе — действительно изолированы, отчуждает нас от Природы и от своих же братьев-людей, тем самым ослабляя нас. Для восстановления нашей человечности в полной мере мы должны снова обрести свой опыт связности, единства со всей паутиной жизни. Это воссоединение, religioпо латыни, есть самая суть духовных основ глубокой экологии. Примечания к главе 12 l. Maturana(1970),MaturanaandVarela(1987),Maturana(1988). Maturana and Varela(1987), pp. 193-4. Умберто Матурана, частная беседа, 1985. См. Maturana and Varela(1987), pp. 212ff. Там же, р. 215. См. Приложение, с. 327–329. Maturana and Varela(1987), p. 234. Там же, р. 245. Там же, р. 244. См. Сарга(1982),р. 302. См. Сарга(1975),р. 88. Varela(1995). См. Сарга(1982),р. 178. См. выше, с. 278. См. Varela et al.(1991), pp. 217ff. См. Capra(1975), pp. 93ff. Cm. Varela et al.(1991), pp. 59ff. Там же, p. 143. Margulis and Sagan(1995), p. 26. Примечания:1 Цитируется по Judson (1979), pp. 209, 220. |
|
||