|
||||
|
|
Глава четвертая СОСТОЯНИЕ СОБАК ПРИ ВТОРОМ СПОСОБЕ ТРЕНИРОВКИ Большой процент суток опытов с аномальным ритмом выделений (57,3%) от общего числа суток, возникновение других неблагоприятных сдвигов, тот факт, что только одна собака оказалась по-настоящему пригодной к длительному пребыванию в кабине и т. д. — все заставляло признать первый постепенный способ тренировки способом, имеющим существенные недостатки. Такой вывод требовал разработки иных методов тренировки. Предлагаемый на следующем этапе исследований второй способ тренировки устранял предварительное приучение собак к условиям малой кабины: животных сразу фиксировали на длительный срок. Была сделана попытка группировки полученных данных по периодам. В качестве основного показателя, по которому проводилось такое разделение, был выбран сон. Начало периода нормализации определялось с момента, когда собаки три дня подряд спали относительно крепко и продолжительно. Предполагалось, что хороший сон способствовал улучшению состояния бодрствования и являлся, таким образом, необходимым фоном для восстановления всех функций. Анализ данных о сне показал, что у собак (Тьмы и Луги) переходный период продолжался только 2 суток, у Угры и Клязьмы — 4–5, у Волхова — 6, а состояние сна Тосны не улучшалось. Вегетативные показатели У большинства собак, как и при первом способе тренировки, в переходный период наблюдались изменения частоты пульса и дыхания. У некоторых животных (например, Луги) в среднем за сутки по сравнению с фоновыми данными они снижались: пульс с 76 ударов в минуту (при сигме 1,27) уменьшался до 72 (при сигме 3,31), частота циклов дыхания — с 12 (при сигме 1,16) до 10 (при сигме 2,41). У других собак изменения были более значительными. У Волхова, например, пульс учащался с 72 (при сигме 2,8) до 83 (при сигме 3,7), частота циклов дыхания — с 16 (при сигме 2,17) до 23 (при сигме 4,35). В отдельные отрезки времени регистрировались задержки дыхания (по 3–5 сек.). На стадии выдоха в переходный период у многих собак, так же как и при тренировке по первому способу, возникали стоны и звуки, производимые в болезненном состоянии. Средние величины числа пульсовых ударов и числа дыхательных движений в переходный период в одну минуту поздним вечером и ночью у многих животных были более учащенными, чем в другие часы суток. Температура тела также ночью бывала на несколько градусов выше (на 0,11–0,22°). У Тосны учащенный на 15–20 ударов пульс, дыхание убыстренное на 3–5 циклов и повышенная на 0,3° температура регистрировались даже в часы глубокой ночи (22–4 час), что говорит о значительных нарушениях вегетатики этой собаки. В ходе опытов по тренировке, т. е. в период нормализации, наблюдались меньшие отклонения величин пульса, числа дыхательных движений и температуры тела. Совершенно исчезали болезненные звуки, сопровождающие дыхание. Исключение составляли вегетативные показатели у собаки Тосны, отличающиеся от нормы. У животного наблюдалась одышка и изменения сердечных ритмов. В период нормализации при ухудшении многих функций также наблюдались повторные увеличения частоты пульса и циклов дыхательных движений, иногда возобновлялись стонущие звуки. Величины этих сдвигов при вторичном их появлении были меньше, чем в переходный период, а при их возникновении в третий раз они были еще менее выражены. Так же как и в тренировке по первому способу, подобные явления были приурочены к актам мочеиспускания и дефекации, а их величины колебались в основном в зависимости от нарушений в этой системе. Все эти данные говорят об изменениях в переходный период вегетативных процессов и относительной их нормализации в ходе дальнейших опытов. Степень изменений была меньшей, чем в опытах по первому способу тренировки. Пищевые рефлексы. Вес В переходный период собаки ели и пили плохо, мало или совсем не реагировали на вид еды. Взвешивание показало несколько меньшее, чем при первом способе тренировки, снижение веса животных (в среднем на 0,26 кг). В начале периода нормализации животные съедали около 10–15% суточного рациона. Начиная примерно с 4–8-х суток аппетит резко улучшался. В конце экспериментов обнаруживалось общее увеличение веса (в среднем на 0,31 кг). Выделения В начале тренировки, так же как и при первом способе, отмечались задержки испражнений: моча впервые появлялась только через 20–35 час. Однако в этих экспериментах в противоположность первому способу тренировки не было обнаружено задержек мочеиспускания по 40 час. и более. Этих актов дольше всего не было у собак Клязьмы и Луги, у Клязьмы затем ритм мочеиспускания быстро нормализовался. Второй раз моча и кал у всех животных появлялись гораздо скорее. Третий и четвертый, т. е. при последующих мочеиспусканиях, интервалы между выделениями были почти нормальными и превышали 7 час. только в том случае, если разделялись ночными часами, когда и в норме эти акты осуществлялись значительно реже. Все эти материалы отражены в табл. 9. Таблица 9. Появление актов мочеиспускания при вторичном способе тренировки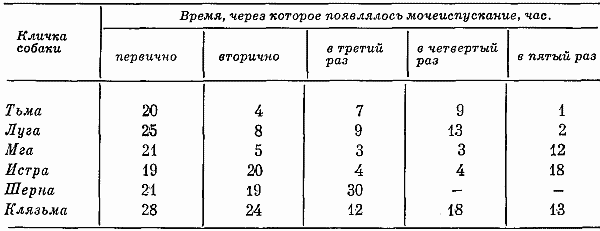 Таблица 10. Частота появления актов мочеиспускания в период тренировки по второму способу Таблица 10. Частота появления актов мочеиспускания в период тренировки по второму способу 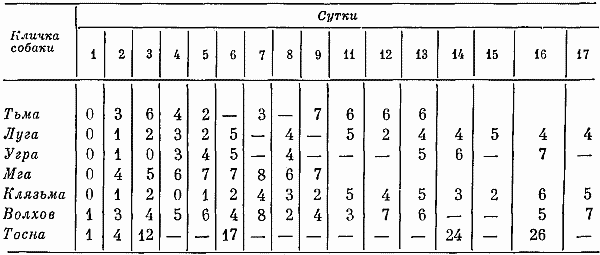 По окончании переходного периода у двух из шести собак сразу установился правильный ритм выделений: мочеиспускание возникало не менее 3 раз в сутки (табл. 10). У одной из собак в 5-е сутки и даже 12-е сутки наблюдался олигоурезис (в виде двухразового выделения мочи), в остальные сутки ритм выделений был нормальным. У другой случаи олигоурезиса регистрировались в 9, 10 и 15-е сутки опыта. Олигоурезис выражался в двухразовом появлении мочеиспускания и только в 3% случаев возникала более тяжелая его форма — одноразовое мочеиспускание в сутки (табл. 10). Нарушения регистрировались в 27% случаев от общего числа суток. Все это говорит о том, что нормализация выделительных процессов по второму способу тренировки происходила менее болезненно, чем по первому. У Тосны, как и по другим показателям, отмечались нарушения ритма мочеиспускания на протяжении всего эксперимента: начиная с 3-х суток, регистрировался палакиурезис. Количество мочеиспусканий за сутки увеличивалось, моча стала появляться в разное время суток, закон соотношения дневного и ночного диуреза нарушался. Сон В начале опытов по второму способу тренировки животные спали мало (иногда только 3–4 часа в сутки) и плохо. Вместо сна часто регистрировался полусон, собаки не находили удобных поз, двигались, пробуждались от слабых раздражений. Данные актограммы во время сна (табл. 11) указывали на неспокойное состояние животных. Число всплесков писчика в среднем было в 3,9 раза большим, чем при 7-часовых опытах, но меньшим, чем в предыдущей серии экспериментов. Таблица 11. Число двигательных реакций за час сна при тренировке по второму способу (по данным актограммы)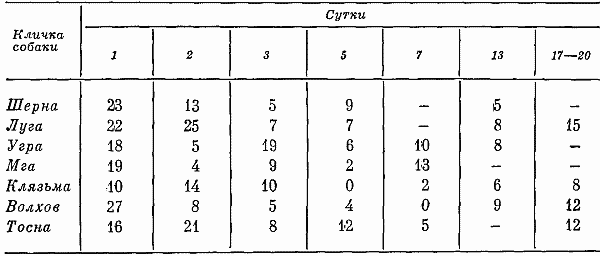 В начале опытов 2–3 суток существенного уменьшения сна сменялись сутками, когда животные спали относительно долго и крепко, потом сон нарушался снова. Во второй половине опытов он нормализовался почти у всех собак. Плохо на протяжении всего опыта спала Тосна: полученные по этой собаке данные говорили о том, что в среднем за сутки она находилась в сонном и полусонном состоянии только 3,3 часа. Интересно отметить, что у всех собак (кроме Тосны) либо отсутствовали, либо были минимальными нарушения сна в период глубокой ночи (22–4 час). То же было и при тренировке по первому способу. Значит, переход к условиям жизнедеятельности в кабине у основной массы животных не разрушал глубоко укрепившихся явлений суточной ритмики. С 4–6-х суток в период нормализации отмечалось превалирование сонных состояний над бодрствованием: животные спали в среднем почти 12 час. в сутки. Более продолжительным сон становился в ранние утренние часы, собаки начинали скорее засыпать вечером, у большинства животных появились случаи дневного сна. Подобные явления наблюдались во время 7-часовых опытов и, следовательно, были нормальными для условий кабины. Поведение Позные состояния. В переходный период животные 68% времени по данным наблюдения находились в активных позах. Расслабленные положения тела регистрировались больше глубокой ночью. Иногда днем наблюдалось возникновение сидячих пассивных поз, что объяснялось значительным ухудшением общего состояния собак. Тосна в основном стояла. В позах покоя она находилась только 16% времени, т. е. менее 4 час, пассивные сидячие позы были для собаки особенно характерным явлением. В ходе дальнейших экспериментов при нормализации состояния собак соотношение поз покоя и действия менялось. Собаки стали сидеть и лежать 58% времени, что в данных условиях было нормальным. Как показали материалы длительных 7-часовых опытов, именно так влияла на собак спокойная обстановка, бедная внешними воздействиями. Двигательное поведение. Так же как и позные состояния, двигательное поведение в основном изменялось по закономерностям, установленным при анализе материалов первой серии экспериментов. В нем сказывались две противоречивые тенденции, одна из которых приводила к сокращению двигательных образований, а другая к их развитию. Пребывание собак в кабине делало для них многие раздражители индифферентными, приводило к уменьшению ориентировочных рефлексов и двигательной деятельности. С другой стороны, развивались двигательные компенсаторные реакции, предохраняющие организм и психику подвижных животных от неблагоприятных сдвигов. У некоторых собак в переходный период двигательное поведение по сравнению с нормой существенно не изменялось, многие проявляли двигательную активность, значительно большую, чем в первой серии экспериментов; другие, наоборот, меньшую: за час бодрствования по данным актограммы одни собаки в среднем производили 203 движения (основной формой активности в переходный период было двигательное беспокойство), а животные, склонные к резкому уменьшению числа движений, — 21 движение. В период 5–7-х суток собаки из группы усиленно двигающихся в среднем производили 83 движения за час до бодрствования, в период 9–12-х и 16–18-х суток — 91 и 104 движения. Группа малоподвижных собак стала двигаться больше: соответственно 38 и 49 движений. Несмотря на то что общее количество движений почти не изменилось, существенно изменился их состав. Двигательное беспокойство, как наиболее элементарная форма таких реакций, почти не возникало. Потребность в движениях реализовалась в виде ритмичных или аритмичных подскоков на месте, раскачиваний, топтаний, смены поз, потягиваний, игры с кормушкой и т. д. У собак часто появлялись такие реагирования на слабые или на средней силы раздражители, какие вызываются сильными сигналами, т. е. утрированные формы двигательных рефлексов (табл. 12). Таблица 12. Изменение (%) в ходе тренировки по второму способу количества движений, возникающих в ответ на внутренние раздражители (по данным сплошного наблюдения, 100% — общее число движений)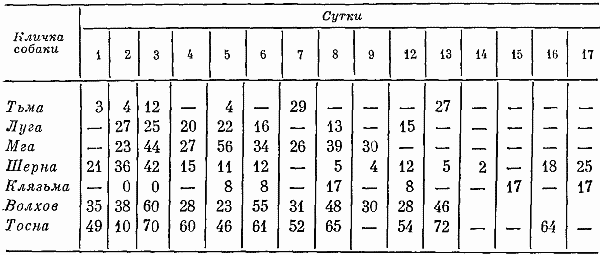 Следовало также отметить развитие у собак, по мере их привыкания к кабине, злобности. О подобном явлении уже говорилось в предыдущих опытах. Отчетливое возрастание во время тренировок злобности почти у всех животных позволяло говорить о ней как о закономерности. Известно, что злобность быстро развивается у животных, посаженных на цепь. Звуковые реакции. При анализе пассивных голосовых реакций были обнаружены те же закономерности, что и при тренировке по первому способу. В самые первые часы опытов, когда собаки еще не испытывали трудности воздействия экспериментов, они не производили звуков. Затем при ухудшении их состояния голосовые реакции получили болезненное выражение (стоны, жалобные скуления). С 4–7-х суток, т. е. в начале периода нормализации, стали возникать лай, визг и рычание, что свидетельствовало о нормализации поведения и об улучшении общего эмоционального состояния животных. По мере продолжения экспериментов звуки, как и при тренировке по первому способу, становились более длительными и разнообразными по своей тональности, форме и другим характеристикам. Так же регистрировались развитые формы феномена прислушивания. Сравнение и анализ данных наблюдений и материалов магнитофонных записей по звуковым реакциям собак при первом и втором способе тренировки говорили о том, что в последнем случае у собак активные голосовые реакции в целом развивались быстрее и носили еще более выраженный характер. А скуление, начиная с 3-х суток, не регистрировалось. В последней трети опытов по второму способу активных звуковых реакций было больше, чем по первому, повторное возникновение скуления появлялось несколько позже. Поведение двух собак (Тосны и Волхова) этой серии требовало особого рассмотрения. Обеих характеризовала чрезмерная двигательная активность и ее неизменность, несмотря на удлинение времени пребывания в опыте. Тосна двигалась 18 час. в сутки (15 из них весьма интенсивно), Волхов — 16 час, что вначале служило причиной их объединения в одну группу. Однако затем были выявлены многие различия в поведении этих собак. Двигаясь, Тосна уставала: интенсивные реакции сменялись неподвижностью в лежачих, а иногда в пассивных сидячих позах. Нос ее часто был теплым и сухим, шерсть потеряла блеск, была запачкана мочой и калом. Отсутствие реакций на внешние раздражители говорило об ухудшении эмоционального состояния. Все эти факты свидетельствовали о существенных сдвигах в функциональном состоянии собаки. Очевидно, она представляла собою животное со многими, как и Ночка, видами патологических реакций, но с другими, нежели та, формами выражения этих реакций в поведении: двигательное беспокойство у Тосны и малоподвижность у Ночки. У Волхова процент активности в ответ на внешние раздражения был выше, чем у других собак, сохранялись адекватные отношения между силой раздражения и реакцией, быстро возникали положительные эмоции. Собака хорошо выглядела: шерсть блестела, нос был холодным и мокрым. В связи с тем, что Волхов, судя по характеру его высшей нервной деятельности, относился к числу животных с сильными процессами возбуждения и торможения, можно думать, что его активность была проявлением сложившихся и поэтому прочных форм поведения. Высшая нервная деятельность За 20–25 дней до начала опытов была сделана попытка охарактеризовать основные особенности нервных процессов каждой подопытной собаки, для чего проводились обследования с помощью проб, подробно описанных на с. 90 этой книги. В силу обстоятельств, изложенных там же, выводы, вытекающие из полученных материалов, носили предположительный характер. У Тьмы, Мги и Шерны уменьшение интервалов между положительными раздражителями при пробах для определения силы возбудительного процесса не сопровождалось ни запредельным торможением, ни существенным изменением латентных периодов, ни появлением большого числа межсигнальных реакций. Значит, нервная система собак выдерживала длительное концентрированное возбуждение. Дифференцировки при удлинении действия тормозного раздражителя до 15 сек. и в большинстве случаев до 20 сек. сохранялись. Пробы для испытания баланса возбудительного и тормозного процесса также обнаруживали высокие показатели уравновешенности этих процессов. У Шерны возбуждение несколько преобладало над торможением, о чем говорили данные во время пробы с удлинением интервала перед тормозным раздражителем. При исследовании высшей нервной деятельности Истры и Луги были получены факты, затрудняющие возможность характеристики основных нервных процессов коры мозга этих животных. Истра при пробе на силу возбуждения не реагировала на последние положительные сигналы, сильно изменялся у нее латентный период реакций, т. е. по сравнению с собаками, о которых говорилось выше, регистрировалась относительная слабость возбудительного процесса. Луга в пробе на возбуждение дала высокие показатели, а при пробах на торможение — выпадение реакций на положительные раздражители при четких реагированиях на торможение. Очевидно, относительно меньшая сила возбудительного процесса у этой собаки была связана с влияниями со стороны сильного тормозного процесса. После дифференцировок при перемежающейся подаче тормозных и положительных раздражителей — проба на баланс нервных процессов — эта собака часто не реагировала на положительные сигналы. То же имело место после одной из двух сдвоенных дифференцировок в пробе с увеличением интервалов между раздражителями. Число межсигнальных реакций у этого животного было небольшим. Волхов при пробе с подачей 20 раздражителей с укороченным интервалом после шестнадцатикратного реагирования на положительные сигналы не отреагировал на два раздражителя. Животное выдерживало удлинение дифференцировочных раздражителей средней и большей интенсивности до 15 сек. (регистрировались межсигнальные реакции), при удлинении тормозных раздражителей до 20 сек. дифференцировки срывались. Все эти факты заставляли предполагать, что функции нервной системы Волхова, характеризуясь в основном сильными процессами возбуждения и торможения, в то же время определялись некоторым превалированием возбудительных процессов. Интересно отметить, что такое заключение в какой-то степени согласовывалось с поведением этой собаки, характеризующимся такими признаками, которые позволяли охарактеризовать это поведение как возбужденное. У Клязьмы проба для характеристики возбудительного процесса (20-разовое применение положительных раздражителей с укороченными интервалами между ними) с самого начала сопровождалась большим количеством межсигнальных реакций. С 6-го до 8-го и с 10-го до 13-го раздражители реакций на положительные сигналы выпали, а с 15-го исчезли до конца обследования, что говорило о появлении запредельного торможения. Процесс торможения этой собаки, судя по данным, полученным при соответствующих пробах, также определялся многими показателями слабости: отмечались частые срывы дифференцировок, были сорваны все реакции на тормозные раздражители большой интенсивности и при удлинении их действия. При испытании баланса возбудительного и тормозного процессов создавалось впечатление о превалировании в реакциях нервной системы Клязьмы тормозных функций (при анализе полученных данных показатели, указывающие на ухудшение тормозного процесса, встречались в 63% случаев, а возбудительного — в 37%). У Тосны слабость возбудительного процесса сочеталась с еще большей слабостью тормозного. При испытании силы раздражения была получена картина, напоминающая таковую у Клязьмы. При пробе на тормозной процесс оказались сорваны все дифференцировки. Их срыв часто наблюдался и при других пробах. Относительная слабость процесса торможения, очевидно, определяла превалирование у Тосны возбудительного процесса. До начала экспериментов условные рефлексы всех собак обладали большой прочностью: собаки реагировали на все положительные раздражители с латентным периодом 1–2 сек., реакции на дифференцировочные раздражители отсутствовали, межсигнальные регистрировались в небольшом числе. Исследование высшей нервной деятельности животных во время 20-суточных экспериментов проводилось в 1–2,8–13 и в 16–19-е сутки опытов. Конкретные данные этих исследований помещены в табл. 13. В 1–2-е сутки опытов реакции нервной системы всех собак оказались существенно изменены. У трех первых животных были случаи отсутствия реагирований на положительные раздражители, несущественно изменялся латентный период реакций на положительные раздражители. Дифференцировки оказались сорванными, собаки реагировали даже на тормозные раздражители в виде света средней интенсивности. Регистрировались межсигнальные реакции. Таблица 13. Число положительных условных реакций дифференцировок и межсигнальных реакций у собак во время 20-суточных опытов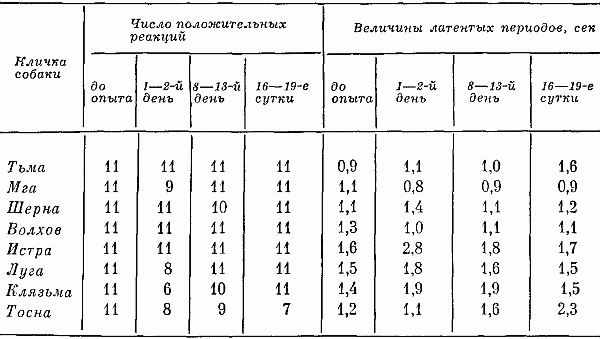 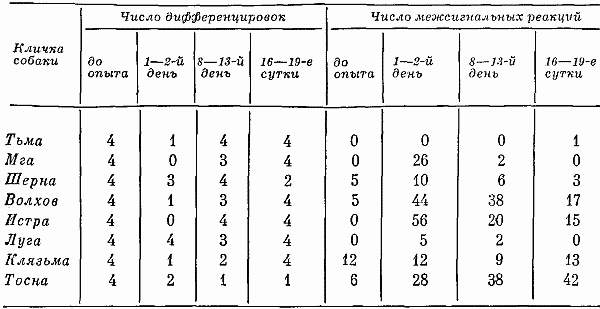 У двух следующих животных также регистрировались срывы дифференцировок на раздражители большой и средней интенсивности, много межсигнальных реакций (44 и 56). У Истры отмечалось значительное увеличение латентного периода реакций. Луга не реагировала на три положительных сигнала, латентный период ее реакций оказался увеличенным. Однако дифференцировки этим животным были выдержаны, число межсигнальных реакций минимально. У Клязьмы и Тосны нарушения высшей нервной деятельности оказались особенно значительными: они реагировали не на все положительные сигналы, дифференцировки в преобладающем большинстве случаев были сорваны, отмечалось много (28) межсигнальных реакций. Эти данные показали, что ухудшение в переходный период многих показателей вегетативных функций животных, показателей их поведения и психических реакций сопровождалось различной степенью функциональных сдвигов в нервной системе. Направление этих сдвигов было не однозначным и не обнаруживало прямой связи между поведением и изменением функций нервной системы. Так, например, Тьма — животное с сильными процессами возбуждения и торможения — вела себя на всем протяжении опыта относительно спокойно. И в этом отношении она была похожа на животное со слабыми процессами возбуждения и торможения (на Клязьму), у которого также не наблюдалось выраженных форм активности. Обращает на себя внимание и тот факт, что при обследовании высшей нервной деятельности в конце 1-х и во 2-е сутки у обеих собак наблюдались одинаковые сдвиги. Волхов и Тосна характеризовались значительной двигательной активностью, несмотря на то что их нервные процессы определялись различными исходными данными. Отсутствие такой связи объяснялось многими причинами. Условия опыта по-разному влияли на животных в зависимости от индивидуальных особенностей их организма и их нервных процессов. Для одних собак сила таких условий была значительной, для других — менее существенной. Свою коррекцию в эти взаимоотношения вносили различные виды поведения, связанные с характерологическими особенностями животных, и неодинаковая степень угнетенного психического состояния. Собаки с сильными нервными процессами не обязательно реагировали на ухудшение условий своей жизнедеятельности наилучшим образом. Например, у Шерны — собаки с сильными нервными процессами менее, чем у других животных, развивались компенсаторные двигательные и звуковые реакции. Развитые формы эти реакции приобрели только в конце эксперимента. Обследование высшей нервной деятельности собак в переходный период опытов говорило о существенном ухудшении реакций нервной системы животных. В основном при этом страдал активный процесс торможения (дифференцировки оказались сохраненными в 37% случаев и полностью — только у одной из восьми собак). Некоторые факты позволяли также предполагать ослабление или даже истощение возбудительных процессов (у Клязьмы и Тосны). Отсутствие у Клязьмы и Тосны в 19% случаев реакций на положительные раздражители, увеличение латентного периода рефлексов заставляли предполагать ослабление возбудительных процессов. Тем не менее ослабление процессов активного торможения в коре мозга этих собак, очевидно, способствовало преобладанию процессов возбуждения. В 8–13-е сутки в высшей нервной деятельности собак регистрировались более благоприятные сдвиги, чем ее показатели в начале эксперимента (см. табл. 12). Все собаки в целом реагировали на 97% подаваемых положительных сигналов. Дифференцировки не были нарушены в 75% случаев подачи дифференцировочных раздражителей. У Мги, Волхова и Луги возникали реакции на тормозной раздражитель большой интенсивности, что говорило о незначительных нарушениях в области условного тормозного процесса коры мозга. У Волхова также отмечалось появление многих межсигнальных реакций. Более существенные изменения показателей нервной системы регистрировались у Клязьмы и Тосны. Собаки реагировали не на все положительные раздражители, латентный период их рефлексов был существенно замедлен, дифференцировки сорваны в пяти из восьми случаев. В конце опытов обследования высшей нервной деятельности (см. табл. 13) наблюдалось дальнейшее улучшение функций нервной системы у преобладающего числа собак и резкое их ухудшение у одного животного (Тосны). Большой интерес в этот период представляли материалы многократных обследований собаки Шерны. Несвойственные для этого животного ошибки при воспроизведении стереотипа и выработанных условных рефлексов в конце опыта были закономерным явлением. Они повторялись в различных соотношениях при специально проведенных исследованиях и показали, что срывы дифференцировок и другие отклонения от выработанного стереотипа реакций являлись результатом растормаживания, возникавшего у этой собаки под влиянием внешних воздействий и собственных двигательных или голосовых реакций: собака, например, до сигнала или в момент его появления начинала топтаться на одном месте или скулить, и это обусловливало появление ошибочных реакций. Растормаживанием также, очевидно, объяснялись многие межсигнальные реакции и других собак (например, у Волхова и Истры). На 2-й и 3-й день после окончания опытов высшая нервная деятельность животных оказалась нормальной. Собаки реагировали на 100% положительных сигналов с латентным периодом до 1,5 сек. и на 94% тормозных, число межсигнальных реакций было минимальным. Сбор характеристик по двигательному и звуковому поведению собак, по их эмоциональному и психическому состоянию, а также нервной деятельности позволили сопоставить эти факты на материале двух серий экспериментов, т. е. по 22 собакам, исключая животных с патологическими формами сдвигов. В переходный период двигательная деятельность животных изменялась в разных направлениях, фиксировалось как ее увеличение, так и затаивание; в области звуков отмечались стоны, скуление и вой. Эмоциональные пробы указывали на изменения эмоций, заключающихся в появлении страха гипертрофированных форм. Обеднения положительных эмоций на этом этапе не отмечалось. Данные, полученные по изучению высшей нервной деятельности, также говорили о многих функциональных сдвигах. В ходе дальнейшего опыта по мере привыкания к его условиям во всех системах организма и психики животных происходили изменения, в основном направленные на приспособление к обстановке. В области поведения это уменьшение движений у большинства неумеренно много двигающихся собак и увеличение двигательных реакций у малоподвижных животных, резкие сокращения скулящих звуков, а затем появление компенсаторных двигательных и звуковых образований. В это же время улучшались эмоциональные и психические реакции, нервная система собак правильно реагировала на большинство положительных и дифференцированных сигналов. Срывы дифференцировок в основном наблюдались при раздражителях большой интенсивности, число межсигнальных реакций сокращалось. В последней трети опытов реакции поведения животных существенно, по сравнению с предыдущим этапом, не изменялись. Однако были и признаки, свидетельствующие о значительном его ухудшении, — возрождение и нарастание скуления. Регистрировалось относительное благополучие в области функций организма (выделительных и других процессов). Реакции поведения, по сравнению с предыдущим этапом, так же значительно не изменялись. Собаки живо реагировали на все изменения в экспериментальной обстановке и, казалось, находились в хорошем психическом состоянии. Но появилось и стало развиваться скуление. В эмоциональных пробах при отсутствии гипертрофированных видов страха, характерных для переходного периода, регистрировалось некоторое ослабление положительных эмоций. Материалы по наблюдению за высшей нервной деятельностью говорили об отсутствии отклонений в функциях нервной системы. Все сказанное позволяло сделать вывод о взаимосвязи в данных опытах состояния организма собак, их поведения, психики и высшей нервной деятельности. Очевидно, в необычных экстремальных условиях такая взаимосвязь проявляется с большей наглядностью. Обсуждение полученных результатов Материалы, приведенные выше и характеризующие первые дни многосуточной фиксации, говорили о неблагоприятных сдвигах в состоянии собак: у животных нарушался ритм выделений, они переставали есть, теряли в весе, начинали плохо и мало спать, у них ослаблялся активный процесс торможения, изменялось поведение, появлялись пассивные звуки. Выраженность таких сдвигов в переходный период, судя по некоторым показателям, была несколько большей, чем в период тренировки по первому способу (по вегетативным показателям, пищевым реакциям, поведению), так как на организм животных, кроме условий, характерных для тренировки по первому способу, действовали факторы, связанные с его первичным пребыванием в кабине. Изменения функций других систем и, главное, выделительной были меньшими. Процесс нормализации по второму способу в основном осуществлялся быстрее и достигал более полного выражения. Анализ неблагоприятных изменений, происходящих у многих животных в переходный период, позволял говорить о их однонаправленности, несмотря на неодинаковые исходные данные. Такая однонаправленность сдвигов предполагала общие причины, лежащие в основе регистрируемых показателей. Очевидно, ими были, как и в предыдущей серии опытов, ломка стереотипа врожденных и выработанных рефлексов ритма выделений, а также резкое сокращение возможности двигаться. Животные должны были преодолеть рефлексы, тормозящие в маленьком жизненном пространстве акты опорожнения мочевого пузыря и прямой кишки. Такое преодоление при тренировке по второму способу возникало быстрее и менее болезненно. Анализ материалов наблюдений за высшей нервной деятельностью показал, что у животных с сильной нервной системой такая перестройка проходила тяжело, но без патологических изменений. Проявлением невротических реакций у одной собаки (Тосны) можно было объяснить резкие неблагоприятные сдвиги в области ритмики, потерю веса, многие поведенческие реакции. Можно высказать предположение, что из двух задач, с которыми сталкивался организм собак, впервые попавших в камеру, а именно с необходимостью отправления выделений и необходимостью находиться в условиях весьма ограниченных движений, первая задача оказалась более трудноразрешимой. К.М. Быков (1944) и В.Н. Черниговский (1960) являлись авторами концепции о взаимосвязи локомоторной деятельности с функциями внутренних органов. По данным, полученным в этих экспериментах, сокращение двигательной активности отрицательно влияло на общее состояние собак. О большом биологическом значении потребности в движениях свидетельствовало двигательное беспокойство и другие уродливые формы проявления в кабине двигательных реакций. Очевидно, отсутствие интенсивной деятельности также было условием, осложняющим отправления выделительной системы. В том случае, когда животные сводили двигательную активность до минимума, у них развивались неблагоприятные явления, и, наоборот, двигательная активность как бы предохраняла животных от этих явлений, о чем говорило относительно хорошее состояние много двигающихся Ветерка, Урагана, Луги, Волхова. Исключение составляла Тосна, двигательная активность которой, организуемая самим животным, не спасала его от развития патологического состояния. Изучение тренировки показало, что двигательное и голосовое беспокойство собак в начале экспериментов было связано с их плохим состоянием, что подтверждалось многими фактами. Это позволяло рассматривать двигательное и голосовое беспокойство собак в начале тренировки как показатели резкого ухудшения состояния. Затем в ходе процесса нормализации собаки становились спокойнее. Еще через некоторое время двигательная и голосовая активность приобретала прямо противоположное значение. Теперь увеличение количества движений и активные звуковые реакции говорили о том, что собаки приспособились к условиям кабины, выработав компенсаторные реакции. Увеличение числа движений и их интенсивности в этом случае было положительным диагностическим признаком состояния животных. Количество движений, возникающих в качестве компенсаторных двигательных образований, очевидно, по своей энергетической стоимости было значительно меньше, чем в нормальных условиях. Е.А. Коваленко, В.Л. Попков с сотрудниками (1971) на основании данных потребления кислорода и выделения углекислоты сделали заключение о существенном снижении газообмена у животных к концу первого месяца. На основании полученных данных можно определить виды реакций, компенсирующих недостаток движений. Это двигательное беспокойство — большое количество немотивированных хаотичных движений с элементами лишних реакций. Значительное место занимали движения, возникающие в силу внутренней потребности двигаться. У одних собак они имели самые разнообразные формы, отличались от двигательного беспокойства большей упорядоченностью и упорностью, у других такие движения выражались в стандартных реакциях. Часто разнообразные действия на глазах экспериментатора стандартизировались и затем осуществлялись преимущественно в такой форме. Как компенсаторные движения также можно рассматривать такие двигательные образования, как, например, потягивание, отряхивание, смену поз. Все эти движения производились животными, находящимися в кабине, чаще, чем в обычных условиях, и с целью подвигаться, а не прямого их назначения. Еще одним видом реакций, компенсирующих недостаток движений, являлись утрированные виды реагирования на внешние раздражители. Четко прослеживаемая зависимость в первые сутки опытов некоторых звуков от патологического состояния говорила о том, что эти реакции в переходный период отражали ухудшение состояния организма и, как показали данные предыдущей серии опытов, психического состояния. На стадии нормализации звуки животных, так же как и движения, играли роль факторов, компенсирующих недостающие моменты нормальной жизнедеятельности в кабине. Такими явлениями была бедность обстановки камерного содержания, отсутствие разнообразных предметов, звуков и запахов. Голосовые реакции самих собак в их различной тональности и формах как бы развлекали животных. Высшая нервная деятельность собак и их психическое состояние в ходе опытов претерпевали изменения. В 1-е и 2-е сутки экспериментов нервная деятельность характеризовалась значительными сдвигами, ухудшалось и психическое состояние. По-видимому, это объяснялось двумя причинами: ухудшением общего состояния организма под влиянием новой обстановки его жизнедеятельности, а также новизной условий, тормозящих разные проявления животных. Последнее было особенно важно для собак второй серии экспериментов, которые фиксировались в кабине без предварительного приучения к ней. Затем по мере продолжения опытов наблюдалось улучшение высшей нервной деятельности и психического состояния. В конце 20-суточных опытов регистрировались в основном высокие показатели высшей нервной деятельности и психики животных и только некоторые моменты свидетельствовали о незначительных изменениях психических реакций. Развитие у собак злобности, базирующейся на повышенной раздражительности, косвенно было свидетельством повышения раздражительности нервной системы и ослабления внутреннего торможения. Отбор собак для камерных исследований Изучение тренировки происходило на протяжении длительного времени и предполагало относительно большое число обследуемых систем. Это требовало большой организационной работы, многих коллективных усилий и длительного времени. Естественно вставал вопрос, нельзя ли в практических целях отбора облегчить эту работу. Для этого на основании проведенного и описанного выше изучения состояния собак в ходе тренировки были разработаны и проверены на практике специальные рекомендации, упрощающие решение вопроса о пригодности собак для их использования в длительных камерных опытах. По этим рекомендациям предполагалось исследование некоторых показателей состояния животных до опыта, а именно: определение их веса, количеств водо- и пищепотребления, определение вегетативных процессов (пульса, количества циклов дыхания и температуры тела). Для того, чтобы изучить фоновые данные этих функций, их следовало регистрировать в течение 5–7 час. с интервалами в 30–60 мин. Затем собаки помещались в камеру и наблюдались там в течение 1-х суток. При этом определялась скорость, с которой у них появлялись акты выделения, регистрировались вегетативные показатели, случаи одышки, звуки при дыхании, стоны, с помощью актограммы фиксировалось состояние двигательной сферы, отмечались позы и пассивные голосовые реакции. Животные на этой стадии обследования считались пригодными для длительных опытов в том случае, если мочеиспускание у них появлялось на протяжении 1-х или 2-х суток экспериментов или в первые часы 3-х и если перечисленные выше явления не носили ярко выраженных и патологических форм. После этого собаки оставлялись в кабине еще на 14–16 суток, в продолжение которых регулярно кормились и осматривались ветеринарным врачом. На 15-е или 17-е сутки проводилось повторное изучение показателей выделительной системы, вегетативных процессов и поведения. Животные могли считаться пригодными к длительным опытам в камере, если в ходе опытов теряли менее 20% своего веса; съедали и выпивали более 60% суточного рациона пищи и воды; у них не было нарушений в сфере выделительных процессов, ишурии, палакиурезиса, стонов при мочеиспускании; не было одышки или звуков при дыхании; отсутствовали патологические формы двигательных и голосовых реакций, чрезмерное количество интенсивных движений, вялость, сидячие пассивные положения тела, неряшливость, двигательные отрицательно эмоционально окрашенные звуки. При особых обстоятельствах такая система обследования собак могла быть сокращена еще более. Животные фиксировались в камере на 15–17 суток и их состояние обследовалось в последние 1–2 дня пребывания в кабине. Об их пригодности судили по признакам, изложенным выше. |
|
||