|
||||
|
|
Наследование пассивно-оборонительного поведения (трусости)[8] в связи с типами нервной системы у собак[9] По мысли И. П. Павлова, на Биологической станции в Колтушах в конце 1934 г. начата работа по изучению генетики высшей нервной деятельности собак. И. П. Павлову не удалось довести эти исследования до конца. После его смерти эта работа продолжается под руководством академика Л. А. Орбели. Над изучением генетики высшей нервной деятельности работает в настоящее время в Институте эволюционной физиологии и патологии высшей нервной деятельности им. И. П. Павлова ряд сотрудников. Однако длительность определения типа нервной системы собаки (около 2 лет), с одной стороны, и относительная медленность размножения собак — с другой, а также исключительная сложность изучаемых признаков весьма замедляют проведение анализа. Несмотря на это, в настоящее время уже можно предварительно говорить о некоторых результатах исследования. Задача настоящей работы — установить, возникает и наследуется ли пассивно-оборонительная реакция собак в какой-нибудь зависимости от типа нервной системы или вне всякой зависимости от него. Постановка такого вопроса важна потому, что изучение пассивно-оборонительной реакции тесно переплелось с изучением типов нервной системы. Пассивно-оборонительная реакция впервые была описана в 1925 г. Ю. П. Фроловым (им же введен и этот термин для обозначения данной реакции поведения). И. П. Павлов упоминал уже об этой реакции поведения, называя ее «прирожденный рефлекс рабской покорности». Он рассматривал этот рефлекс как целесообразное проявление организма: «Нарочитая пассивность позы слабейшего, естественно, ведет к падению агрессивной реакции сильнейшего, тогда как хотя бы и бессильное сопротивление слабейшего усиливает раздражительное возбуждение сильнейшего». (Павлов, 1938. С. 348). Описываемая Фроловым собака по кличке Меля обладала значительно выраженной пассивно-оборонительной реакцией: «Не только привязь, но даже вся обстановка комнаты и лаборатории являются для нее как бы гипнотизирующими моментами. Они погружают ее в то состояние, в котором приказы и внушения выполняются беспрекословно» (Фролов, 1925. С. 335). Фролов сближает эту особенность с врожденными рефлексами (инстинктами). Происшедшее в 1924 г. наводнение в Ленинграде дало материал для изучения влияния на пассивно-оборонительную реакцию такого сильного раздражителя, как это стихийное бедствие. Сперанским (1927) была описана собака с пассивно-оборонительной реакцией, резко изменившая свою условно-рефлекторную деятельность после действия на нее этого необычайного раздражителя. В результате возникшего невротического состояния произошло торможение всей условно-рефлекторной деятельности собаки. Величина условных рефлексов упала до нуля. Потребовалось много дней для того, чтобы восстановилось нормальное поведение собаки. Рикман (1928) наблюдал торможение условных рефлексов при применении сильных раздражителей у собак с пассивно-оборонительной реакцией. На основании этих фактов Павлов пришел к выводу, что в основе нормальной боязливости, трусости, а особенно болезненных фобий, лежит простое преобладание физиологического процесса торможения как выражение слабости корковых клеток. Описывая основные типы нервной системы собак, Павлов помещал на крайние полюса, с одной стороны, возбудимых собак, а с другой — тормозных (трусливых). «Возбудимый тип в его высшем проявлении — это большей частью животное агрессивного характера, например, если хозяин, которого они хорошо знают и которому они вполне покоряются, с ними поступит резко, ударит, они могут его укусить, не удержаться. Крайний тормозимый тип выражается в том, что стоит на собаку прикрикнуть, замахнуться, чтобы она поджала хвост, присела, даже помочилась. Это то, что называется трусливое животное» (Павлов, 1938. С. 466–467). Из этой цитаты ясно видно, что в тот период (1927 г.) Павлов противопоставлял возбудимых животных трусливым, считая последних «специалистами торможения». Таким образом, согласно взгляду Павлова и его школы того времени, эти крайние типы являлись диаметрально противоположными, взаимно исключающими, и трусливые собаки считались и описывались как «слабый тормозной тип нервной системы». Однако в дальнейшем Иванов-Смоленский (1932) провел специальное исследование на трусливой собаке и показал, что, несмотря на то что она обладала резко выраженной пассивно-оборонительной реакцией, тем не менее не могла быть отнесена к слабому типу нервной системы. Эта работа впервые показала, что трусливые собаки не обязательно должны быть слабыми. Майоровым (1933) была проведена специальная работа с воспитанием щенят в условиях изоляции. В то время как контрольные собаки тех же пометов, воспитывавшиеся на свободе, не проявляли пассивно-оборонительной реакции, собаки, воспитывавшиеся в условиях изоляции, проявляли ее в резко выраженной степени (как по отношению к людям, так и ко всякой новой обстановке) и, по мнению автора, несмотря на свой более чем двухлетней возраст, сохранили щенячий тип поведения, не вступив ни разу в борьбу за существование с внешними агентами всякого рода. Майоров не считает своих собак принадлежащими к слабому типу нервной системы. Одним из фактов, подтверждающих это, является постепенное ослабление пассивно-оборонительной реакции у «заключенных» собак. Эти опыты указывают на то, что пассивно-оборонительная реакция, очевидно, не обязательно должна быть связана со слабым типом нервной системы. Розенталь (1930), поставив специальные опыты на собаке с пассивно-оборонительной реакцией, относившейся в течение ряда лет к слабому типу, показал, что эта собака принадлежит не к слабому типу нервной системы, а, наоборот, должна быть отнесена к сильному типу. Розенталь предположил, что у его собаки оказался неизжитым «детский рефлекс боязни». Однако, не считая, что пассивно-оборонительный рефлекс обязательно должен быть связан со слабым типом нервной системы, Розенталь полагает, что щенята сильного типа могут с возрастом изжить «детский» рефлекс осторожности, в то время как у щенят слабого типа полная задержка такого рефлекса невозможна, как невозможна она у собак сильного типа нервной системы, но при особенно неблагоприятных условиях воспитания. На основании всех этих фактов И. П. Павлов в последние годы своей жизни отошел от первоначального отождествления трусости и слабости нервной системы: «Все казавшиеся нам трусливыми, т. е. медленно привыкавшие к нашей экспериментальной обстановке собаки, которые также с трудом вырабатывали условные рефлексы (а вся их условнорефлекторная деятельность легко может быть нарушенной от малозначительных новых внешних влияний), огульно относились нами к слабому типу нервной системы. Это повело даже к грубой ошибке, когда я одно время считал таких собак специалистами торможения, т. е. сильными по торможению» (Павлов, 1938. С. 655). Таким образом, Павлов отказался в последние годы своей жизни от отождествления пассивно-оборонительной реакции со слабым типом нервной системы, считая трусость реакцией, которая постоянно маскирует истинную силу нервной системы. Генетическая литература по изучению признаков поведения у собак немногочисленна. Однако есть материал, который указывает, что ряд признаков поведения, и в том числе оборонительные реакции, наследуются. Эти данные охватывают главным образом признаки поведения, связанные, очевидно, с подкорковыми отделами нервной системы. Что касается наших знаний по наследованию реакций поведения, связанных со свойствами головного мозга, то в этом отношении литературные данные особенно незначительны. Однако имеется материал, указывающий на роль наследственных факторов в способности к выработке условных рефлексов у собак и мышей. Остановимся прежде всего на литературе по оборонительным реакциям поведения. Начнем с данных, полученных на собаках. Хамфри и Вернер (Humphrey, 1934) изучали наследственные признаки поведения, и в частности оборонительных реакций, у собак. На основании собственных данных они приходят к выводу, что боязнь сильных тактильных раздражителей и сильных резких звуков является самостоятельным рецессивным наследственным признаком. В своем проявлении и выражении они находятся в зависимости от предшествующей жизни собаки. Крушинский (1938) изучал наследование оборонительного поведения у собак. Согласно его данным, наследственность играет существенную роль в возникновении пассивно-оборонительной реакции у собак. Данные автора свидетельствуют также о возможности наследования пассивно-оборонительной реакции в качестве доминантного признака. Однако автор указывает, что различные условия содержания и воспитания собак, играющие столь большую роль в возникновении данной реакции поведения, заставляют делать подобные выводы с большой осторожностью. В обсуждении полученных результатов указывается как на одну из возможностей наследование не собственно пассивно-оборонительной реакции, а ее слабого типа, и лишь на его фоне развивается пассивно-оборонительное поведение. В этой работе приведен материал по наследованию активно-оборонительной реакции (злобности), который свидетельствует о вероятности наследования злобности в качестве доминантного признака. Проявление и выражение активно- и пассивно-оборонительной реакции находятся, по данным Крушинского, в большой зависимости от степени общей возбудимости собаки: чем выше возбудимость, тем сильнее выражение оборонительного поведения. Комбинация повышенной возбудимости с оборонительными реакциями приводит к значительному усилению выражения последних. Этим дается объяснение гетерозиса в выражении пассивно-оборонительной реакции, который наблюдался при гибридизации гиляцких лаек с немецкими овчарками и собак с волками. Данные гибридологического анализа подтверждаются в этих исследованиях опытами с искусственным повышением возбудимости у трусливых и злобных собак. В результате инъекции кокаина или скармливания гормона щитовидной железы происходит усиление выражения имеющейся у собаки оборонительной реакции, и наоборот, понижение возбудимости, происходящее в результате экстирпации щитовидной железы, приводит к ослаблению этой реакции. Таким образом Крушинский, указывая на значительную роль наследственности в возникновении оборонительных реакций, считает, что они в своем проявлении и выражении зависят от весьма различных условий. Интересны данные по гибридам волков и собак. Относительно этих гибридов у практиков-собаководов имеется совершенно определенное мнение о постоянном проявлении у них резко выраженной трусости, более резкой, чем у волков. Шмидт (Schmidt, 1940) описал поведение гибридов волков с собаками. Он указал на проявление пугливости, боязни новых предметов у гибридов первого поколения, происходящих от скрещивания волка с собакой. Во втором поколении произошло ясное расщепление по признакам поведения. Хамфри и Вернер (Humphry, 1934) указывают на попытку дрессировать гибридов волков с собаками. Дрессировка продвигалась успешно до тех пор, пока гибриды находились на привязи, но как только они оказывались спущенными с привязи, невозможно было заставить их повиноваться. Адамец указывает, что склонность к одичанию, которую можно поставить в известную связь с пассивно-оборонительной реакцией, наиболее характерна, по наблюдениям овцеводов Патагонии, для гибридов овчарок. Эти литературные данные свидетельствуют о том, что наследственность играет значительную роль в возникновении оборонительных реакций у собак. Работы, проведенные с наследованием оборонительных реакций на других животных, находятся в полном согласии с этими данными. В основном эти работы проведены на крысах и мышах. Иеркес, работая на крысах, показал, что такие свойства, как злобность, дикость и трусость, являются наследственными признаками. Кабурн на основании большого материала (1300 мышей) пришел к выводу о наследовании «дикости». Он считает, что этот признак определяется не одним, а несколькими наследственными факторами. Садовникова-Кольцова (1925, 1928), работая с крысами, показала различное поведение в лабиринте животных, происходящих от «диких» и различных линий «ручных» крыс. Даусон (Dawson, 1932) изучал наследование «дикости» у мышей. На большом материале (3376 животных), применяя точную методику учета интенсивности реакции, автор показал, что «дикость» наследуется как доминантный фактор, на выражение которого влияет несколько модификаторов. На других животных наследование этих реакций планомерно не исследовалось. Можно указать лишь на данные Хофманна о том, что горячий злобный темперамент наследуется у лошадей как доминантный признак. Филлипс (Phillips, 1912), скрещивая различные породы уток, пришел к выводу, что пугливость у них обусловливается генетически, однако значительное влияние оказывают также условия содержания. Таким образом, все эти данные указывают на то, что наследственность играет существенную роль в возникновении оборонительных реакций у различных животных. Генетические факторы, очевидно, играют значительную роль в возникновении и других признаков поведения у собак. Наиболее детально исследована роль наследственности в возникновении отдельных реакций поведения у охотничьих собак. Хорошо известно огромное значение дрессировки (натаскивания, нагонки) для успешной работы охотничьих собак. Однако значительная роль принадлежит в этом также и наследственности. Уитней (Whitney, 1932) исследовал наследование гона (trial barking) собак. В результате скрещивания собак, гоняющих зверя по следу с лаем и без лая, в первом поколении рождалось всегда лающее на следы зверя потомство. Однако интонация лая, характерная для гончих, не наследовалась. Во втором поколении рождались как лающие, так и не лающие на следу зверя собаки. Склонность легавых собак делать стойку, т. е. останавливаться перед дичью, привлекала внимание еще Дарвина. Говоря о возникновении этой особенности поведения, он указывал, что раз появились собаки, останавливающиеся перед дичью, что, возможно, явилось остановкой перед броском на добычу, то систематический отбор, который продолжается и теперь, усилил развитие этого признака. В последнее время Мархлевский (Marchlewsky, 1930) пришел к выводу, что резко выраженная способность к стойке, характерная для английского пойнтера, имеет неполное доминирование над менее выраженной тенденцией к ней, характерной для немецкой легавой. Автор указывает, что способность к так называемому «секундированию» (backing — у англичан), которая проявляется в том, что собака делает стойку не только по запаху дичи, но и при виде другой собаки, стоящей на стойке, зависит от особого наследственного фактора, имеющего различное выражение в разных линиях охотничьих собак.[10] Манера разыскивать птицу нижним чутьем, т. е. по следам, и поэтому с головой, опущенной вниз (часто встречающаяся у немецких легавых), является рецессивной по отношению к классическому свойству пойнтеров высоко держать голову во время поиска и использовать воздушные течения для улавливания запахов, исходящих от самой птицы, а не ее следа (Marchlewsky, 1930; Whitney 1932). По Мархлевскому, манера молодых собак преследовать добычу с подлаиванием наподобие гончих (свойство, которое проявляется у немецких легавых) является рецессивной по отношению к «молчаливому» способу охоты, которое характерно для пойнтеров. Согласно данным Уитней, большая склонность некоторых охотничьих собак залезать в воду наследуется как доминантный признак, склонность к охоте за птицами наследуется как не полностью доминантное свойство. Говоря о генетическом анализе охотничьего поведения у собак, надо иметь в виду, что приведенные нами данные еще далеко не позволяют установить истинную картину наследования этих свойств. Говоря о рецессивном или доминантном наследовании того или другого признака поведения, авторы констатировали в основном только степень сходства гибридов с одним из родителей. Все перечисленные выше наследственные признаки поведения связаны, по-видимому, в основном со свойствами подкорковых отделов нервной системы. Что касается данных генетического анализа признаков, связанных со свойствами коры головного мозга, то в литературе таких данных очень немного. Так, Хамфри и Варнер (Humphrey, 1934), производя селекцию по рабочим качествам служебных собак, показали, что селекция улучшила рабочие качества собак той популяции, с которой проводилась работа. Однако дрессировка служебных собак, построенная далеко не на одних свойствах коры, не дает возможности судить по суммарному результату улучшения рабочих качеств, какие свойства нервной системы подвергались в данном случае отбору. Даусон и Кац сообщили предварительные данные по изменчивости способностей к обучаемости у собак. Цель работы — создать основу для возможности дальнейшего изучения генетики поведения. Перед подопытными собаками ставилась задача правильного выбора незапертой двери (одной из четырех). В этих опытах некоторые собаки развивали хорошую систему выбора правильной двери. Всего было изучено 44 собаки нескольких пород. Проведенные опыты показали, что наблюдается большая изменчивость по обучаемости как между отдельными собаками, так и между отдельными породами. Скрещивание с собакой, давшей наиболее высокие показатели обучаемости, увеличило и средние показатели обучаемости ее потомков по сравнению со средними показателями для всей популяции, показав тем самым эффективность селекции по данному свойству. Наиболее детальной работой по изучению роли наследственности в обучаемости, т. е. способности к выработке условных рефлексов, является работа Викари (Vicari, 1929), проведенная на мышах. Была изучена обучаемость японских танцующих мышей (Mus wagneri asiatica) и трех линий обычных мышей (Mus musculus): альбиносов, ослабленных коричневых (dilute brown) и ненормальноглазных (abnormal (x-ray-eyed). Оказалось, что каждая линия мышей имеет характерную для нее кривую обучаемости. Скрещивания, проведенные между представителями этих линий мышей, показали, что кривая быстрой обучаемости линий коричневых мышей доминирует над кривой более медленной обучаемости линии ненормальноглазых мышей и еще более медленной обучаемости альбиносов. Кривая обучаемости последних доминирует, в свою очередь, над кривой обучаемости японских танцующих мышей. На основании расщепления, полученного во втором поколении, автор указывает как на одну из возможностей, что различие в обучаемости между коричневыми и белыми мышами обусловлено монофакториально. Однако не отрицается возможность и более сложной картины наследования. Различие в обучаемости между линией альбиносов и японскими танцующими мышами, представителями двух разных видов, обусловливается наличием нескольких факторов. Большое количество подопытных животных (900 мышей), детальный анализ и обработка материала с несомненностью указывают на большую роль наследственности в способности к обучаемости мышей. Таким образом, из приведенных данных видно, что определенные свойства поведения, связанные с корой головного мозга, могут, очевидно, наследоваться. Можно указать еще на генетический анализ такого общего свойства нервной системы, которое находит свое внешнее выражение в нервно-мускульной активности, как общая возбудимость животного. Адамец (Adametz, 1930) указывает, что повышенная возбудимость, нервозность английского пойнтера не полностью доминирует над флегматичным темпераментом немецкой легавой. Подобные скрещивания практиковались в южных областях Моравии, так как в летнюю жару быстро работающие пойнтеры скоро доходили до изнеможения; получавшиеся же метисы обладали желательным «выравненным темпераментом» и оказывались превосходными рабочими собаками. Крушинский в цитированной выше работе исходил из характеристики повышенно возбудимых собак, сделанной И. П. Павловым. Было показано, что гибриды в количестве 25 экз., полученные от скрещивания повышенно возбудимых немецких овчарок с мало возбудимыми гиляцкими лайками, оказались в первом поколении «или подобно немецким овчаркам сильно возбудимыми, или средне возбудимыми, но все они были возбудимее своих отцов гиляцких лаек». Эти данные говорили о вероятности наследования повышенной возбудимости или в качестве доминантного или не полностью доминантного признака. Андерсон (Anderson, 1939), используя методику учета активности, возбудимости собак при помощи шагомера, указывает, что различные породы имеют весьма различную суточную активность. Можно выделить по этому признаку три группы собак: к наиболее возбудимой, активной группе принадлежат немецкие овчарки, арабские борзые и спаниели; к среднеактивной группе принадлежат английские бульдоги, таксы и пекинские собаки; к группе малоактивных собак принадлежат французские гончие. Были получены следующие потомки: французские гончие ? немецкие овчарки F1 и F2; французские гончие ? арабские борзые F1 и F2; французские гончие ? английские бульдоги; арабские борзые ? пекинские собаки F1 и F2. По возбудимости, активности гибриды первого поколения оказались промежуточными между крайними родительскими группами. Большинство гибридов второго поколения по своим показателям приближались к гибридам первого поколения, однако отдельные особи давали очень высокие и очень низкие цифры, указывая на очевидность происшедшего расщепления. Таким образом, эти совпадающие данные, полученные различными авторами, с несомненностью говорят о том, что повышенная возбудимость, активность собак является в значительной степени наследственным признаком. Эти результаты, полученные на собаках, подтверждаются исследованиями Садовниковой-Кольцовой на крысах. Изучая активность крыс в колесах Гринмана, она показала, что при скрещивании диких крыс, дававших очень высокие показатели суточной активности, с лабораторными крысами, дававшими во много раз более низкие показатели, гибриды первого поколения оказались промежуточными по своей активности. Приведенные нами данные генетического анализа признаков поведения с несомненностью указывают, что ряд свойств поведения животных обусловлен в определенной степени наследственностью. Это относится не только к таким свойствам поведения, как оборонительные реакции или признаки охотничьего поведения собак, но, очевидно, и к определенным свойствам коры головного мозга и более общему свойству нервной системы — степени ее общей возбудимости. На основании изложенного выше можно сказать следующее: пассивно-оборонительная реакция собак зависит в своем возникновении, очевидно, от двух причин — от условий содержания и от наследственности. Данные некоторых авторов (Иванов-Смоленский, 1932; Розенталь, 1930) указывают на то, что собаки с пассивно-оборонительной реакцией не обязательно должны быть слабыми. Таково же было мнение И. П. Павлова в последние годы его жизни. Генетический анализ оборонительного поведения в его зависимости от типологических особенностей собак, проведенный в настоящей работе, можно надеяться, дополнит высказанные И. П. Павловым мысли по этому вопросу. Материал и метод Материалом для данной работы послужили собаки питомника Института эволюционной физиологии и патологии высшей нервной деятельности им. И. П. Павлова. Подавляющее большинство собак воспитывались в идентичных условиях того же питомника. Почти все собаки были беспородными. Оборонительные реакции всех подопытных собак были определены одним и тем же человеком (Крушинским). Типы нервной системы подопытных собак определялись различными работниками (Тимофеевой, Трошихиным, Зевальдом, Колесниковым, Красуским, Плешковым, Болоховским и др.). Так как пассивно-оборонительная реакция собак имеет весьма различное выражение, начиная от самой незначительной, едва уловимой степени и кончая резко выраженной патологической формой, необходима количественная оценка этой реакции. С этой целью была разработана шкала, отображающая наиболее характерные черты поведения пассивно-оборонительных собак. Все собаки по этой шкале делятся на 7 основных групп. Начиная от группы собак с полным отсутствием трусости, даются наиболее характерные черты возрастающей степени пассивно-оборонительной реакции. Пассивно-оборонительное поведение каждой собаки может быть отнесено к одной из степеней выражения трусости по этой шкале, что дает возможность ее количественной оценки. Определение оборонительного поведения всегда производилось в одних и тех же условиях: в вольерах питомника (обычном месте жизни собак), в присутствии хозяина собаки (человека, ухаживающего за ней на питомнике). Раздражителем для выявления пассивно-оборонительной реакции был всегда один и тот же незнакомый для собаки человек (Крушинский), по возможности соблюдавший один и тот же стандарт движений в присутствии испытуемой собаки. Приведем шкалу, по которой производилась оценка пассивно-оборонительной реакции: Тр-0.[11] (совершенно нетрусливые собаки). При приближении к собаке незнакомого человека не проявляет никаких признаков трусости. При заходе в вольер и при внезапном раздражителе (удар в ладоши около самой собаки) не проявляет никаких признаков испуга. Тр-1.[12] При приближении к собаке незнакомого человека не проявляет никаких признаков испуга. При приближении незнакомого человека к самой клетке, в которой сидит собака, не проявляет признаков испуга; при внезапном звуковом раздражителе несколько прижимается, приседая к полу; иногда отбегает на 1–2 шага, но затем тотчас же всякие признаки испуга исчезают. Тр-2. При приближении к собаке незнакомого человека отбегает на несколько шагов. При подзывании незнакомым человеком осторожно подходит к нему, присев на согнутых ногах. При звуковом раздражителе отбегает. Тр-3. При приближении к собаке незнакомого человека отбегает. При подзывании незнакомым человеком не подходит к нему. При подзывании собаки ее хозяином (ухаживающим за нею человеком), который стоит в 2–3 шагах от незнакомого человека, подходит к хозяину. Тр-4. При приближении к собаке незнакомого человека убегает; обычно забивается в угол вольеры. При подзывании незнакомым человеком не подходит. При подзывании хозяином не подходит, однако делает попытки подойти к нему (тянется, делает по направлению к нему 1–2 шага). Тр-5. При приближении к собаке незнакомого человека забивается в угол. В присутствии незнакомого лица на подзывание хозяина не подходит и не делает попыток подойти. После ухода незнакомого человека сразу выходит из угла, в котором пряталась. Тр-6. В присутствии незнакомого человека сидит, забившись в угол. На подзывание хозяином в присутствии незнакомого человека не подходит и не делает попыток подойти. Обычно наблюдается мочеиспускание, иногда дефекация. После ухода незнакомого человека долгое время не выходит из своего убежища. Пассивно-оборонительная реакция у собак впервые проявляется нередко только в годовалом возрасте, а иногда и позднее. Поэтому окончательная характеристика пассивно-оборонительного поведения может быть дана не раньше, чем в полуторагодовалом, а еще лучше — двухгодовалом возрасте. У взрослой собаки оборонительное поведение является константным, сохраняя в течение ряда лет при одинаковых условиях содержания сходную форму своего выражения. В табл. 1 даны результаты определения пассивно-оборонительной реакции у собак в течение двух и трех последующих лет; причем для исключения возможности субъективной оценки исследователь не имел перед собой результатов предыдущего обследования собак. Таблица 1. Результаты обследования собак по признаку пассивно-оборонительной реакции в течение трех лет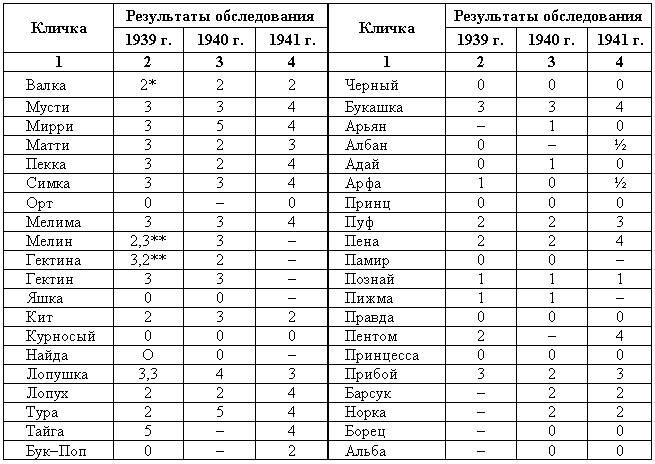 * Всюду опущено обозначение «Тр», а указана только степень трусости. ** Приведены результаты двух обследований в одном и том же году. Коэффициент корреляции между последним определением и предыдущим равен для этой таблицы r= +0,87+0,04 и указывает на хорошее совпадение результатов оценок в двух разных годах определения. Из таблицы видно, во-первых, что собаки, имевшие Тр-0 (нетрусливые), в большинстве случаев проявляют в последующие годы сходную картину поведения. Отклонения в показателях, которые у них наблюдаются, не превышают в большинстве случаев оценки Тр-1; во-вторых, собаки, имеющие оценки Тр-2 и выше, не имеют в большинстве случаев оценки ниже этого показателя. Эти данные показывают, что можно выделить две основные группы собак: с пассивно-оборонительной реакцией и с ее отсутствием. В первую группу входят собаки начиная от Тр-2 и выше, во вторую — собаки группы Тр-0 — Тр-1, так как те следы пассивно-оборонительной реакции, которые проявляются при внезапном звуковом раздражителе у собак группы Тр-1, не являются настоящей пассивно-оборонительной реакцией. Как эти данные, так и высокий коэффициент корреляции между отдельными определениями показывают, что оборонительное поведение является признаком, сохраняющим довольно постоянную картину своего выражения в течение ряда лет. Колебания в результатах отдельных оценок могут зависеть от двух причин: от колебания выражения самого признака (оборонительного поведения) и от методики его учета. Вероятно, эти колебания у обследованных нами собак зависят и от того и от другого. Трудно допустить, чтобы оборонительное поведение совершенно не менялось в течение ряда лет, но в то же время вполне возможно и некоторое расхождение в отдельных оценках при определении трусости даже при одинаковом выражении этого признака. Типы нервной системы определялись по стандарту, принятому в Институте эволюционной физиологии и патологии высшей нервной деятельности им. Павлова.[13] Работа велась в основном по следующим направлениям: 1) определялась сила раздражительного процесса (показатели: быстрота выработки условных рефлексов: изменение условных рефлексов при суточном голодании, влиянии различных доз кофеина и действии сильного раздражителя — трещотки; 2) определялась сила тормозного процесса (показатели: быстрота выработки дифференцировки; изменение условных рефлексов при влиянии различных доз брома, удлинении дифференцировки и сшибке[14] нервных процессов; 3) определялась подвижность нервных процессов (показатели: условно-рефлекторная работа при изменении стереотипа, быстрота переделки положительного рефлекса в отрицательный и отрицательного в положительный; выработка запаздывающего рефлекса). На основании работы с собакой дается характеристика ее нервной системы. Выделяются четыре основных типа в том виде, как они даны И. П. Павловым. Все собаки делятся по силе их нервной системы на особей с сильными и слабыми нервными процессами. У слабых нервная система истощается быстрее и при сильных раздражителях легко впадает в запредельное торможение. В то время как повышение возбудимости коры головного мозга (голодом, определенными дозами кофеина, сильным раздражителем) приводит у сильной собаки к повышению величины условных рефлексов, у слабой собаки происходит их понижение. Сангвиники отличаются от флегматиков подвижностью нервных процессов (сангвиники — хорошая подвижность, флегматики — плохая подвижность). Холерики характеризуются значительным отставанием процесса торможения от процесса раздражения. Так как свойства нервной системы собак обнаруживают бесконечные вариации, приходится обозначать отдельных собак с указанием их отклонения в ту или другую сторону от чистого типа, например: слабый тип нервной системы, показывающий по некоторым тестам вариацию в сторону сильного типа, обозначается как «слабый тип сильной вариации», и наоборот, сангвиник, проявляющий некоторые черты слабости нервной системы, обозначается как «слабый вариант сангвиника»; если у сангвиника несколько отстает тормозной процесс, такая собака обозначается как сангвиник с «несколько отстающим тормозным процессом»; подобные же варианты могут наблюдаться и по свойству подвижности процессов возбуждения и торможения. Фактический материал Весь фактический материал настоящей работы сведен в родословные отдельных семейств собак. В родословных приведены данные оценки оборонительного поведения[15] каждой собаки и результаты типологического анализа. Оборонительное поведение каждой собаки выражено в родословной различной степенью зачернения. Тип собаки, в виде условного обозначения, ставится снизу в родословной под изображением каждой собаки. Семейство № 1 (рис. 1). В скрещивании I этого семейства получены потомки от двух сангвиников — Агути и Томбуша. Практически можно считать, что ни у той, ни у другой собаки пассивно-оборонительной реакции нет, хотя у Агути наблюдались самые незначительные следы этой реакции. Все потомки от этого скрещивания оказались сильными; один из них — Арфа (сангвиник ниже среднего) — проявила самую незначительную пассивно-оборонительную реакцию (Тр-1). Обращает на себя внимание то, что у Агути наблюдалось некоторое отставание тормозного процесса и подвижности. У двух ее потомков (Албан, Адай) отстает подвижность и у двух (Арьян, Адай) — тормозной процесс. В скрещивании II этого семейства получены потомки от Томбуша и его дочери Арфы. Один из потомков этого скрещивания (Арт) проявил значительную пассивно-оборонительную реакцию (Тр-4); остальные потомки или не имеют ее совершенно, или имеют в очень незначительной степени. В общем можно сказать, что в двух этих скрещиваниях подавляющее большинство потомков не проявили или проявили пассивно-оборонительную реакцию в очень незначительной степени. Иная картина наблюдается в скрещивании III этого семейства, в котором получены потомки от скрещивания Томбуша с Пеной. Пена проявляет, несмотря на сильный тип нервной деятельности, ясно выраженную пассивно-оборонительную реакцию (Тр-4). У трех из девяти ее потомков проявилась ясно выраженная пассивно-оборонительная реакция (Тр-3 и Тр-4); у остальных ее потомков ее или нет, или она выражена незначительно. Несмотря на проявление трусости, двое из потомков оказались сильными (Прибой, Пуф). Семейство № 2 (рис. 2). В этом семействе средний сангвиник — Букашка — с значительно выраженной трусостью (Тр-4) скрещивается с очень сильным, совершенно нетрусливым сангвиником Черным, обладающим очень хорошей подвижностью. Все потомки от этого скрещивания проявили пассивно-оборонительную реакцию: трое из них оказались сильной вариацией слабого типа, остальные — сангвиниками сильной и слабой вариации (определение не закончено). В данном скрещивании значительная пассивно-оборонительная реакция комбинируется как со слабым (сильной вариации), так и с сильным типом нервной системы. Как в этом, так и в следующем семействе ясно выступает то, что пассивно-оборонительная реакция легко комбинируется с сильным типом нервной системы. 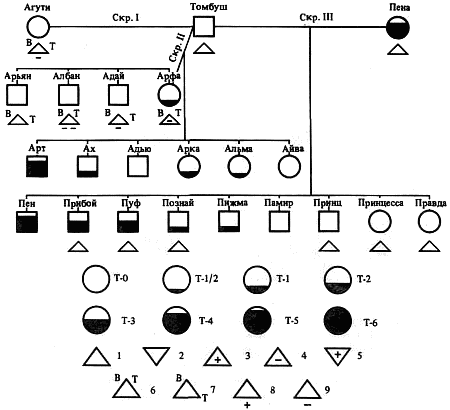 Рис. 1. Семейство № 1 Рис. 1. Семейство № 1 На рис. 1–6: 1 — сильный тип; 2 — слабый тип; 3 — сильный тип, сильная вариация; 4 — сильный тип, слабая вариация; 5 — слабый тип, сильная вариация; 6 — сангвиник; 7 — холерик; 8 — хорошая подвижность нервных процессов; 9 — плохая подвижность нервных процессов. 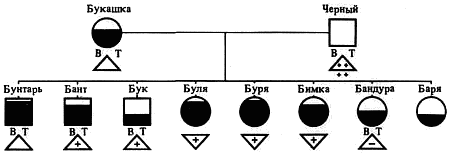 Рис. 2. Семейство № 2 Рис. 2. Семейство № 2 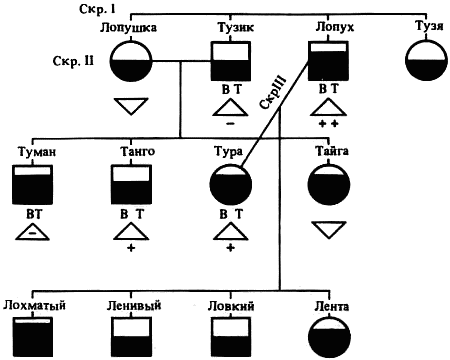 Рис. 3. Семейство № 3 Рис. 3. Семейство № 3 Семейство № 3 (рис. 3). У всех представителей этого семейства в ясной форме выражена пассивно-оборонительная реакция; в то же время большинство из них оказались сильного типа. Две слабые собаки отнюдь не являются более трусливыми, чем сильные представители этого семейства. Семейство № 4 (рис. 4). В этом семействе (скрещивание I) трусливая слабая сука Дедди при скрещивании с нетрусливым сильным кобелем Гектором[16] дала четырех потомков, у которых в той или другой степени была выражена пассивно-оборонительная реакция (Тр-3, Тр-4 и Тр-5). Две самки, полученные от этого скрещивания, оказались слабого типа; оба самца — сангвиники слабой вариации. Скрещивание II двух из этих потомков (Мелина и Мелин) между собой дало помет, в котором три собаки оказались с пассивно-оборонительной реакцией, а у одной этой реакции не было. Эта собака оказалась вместе с тем очень сильным сангвиником. Три других потомка, проявивших пассивно-оборонительную реакцию (Тр-2), оказались сангвиником, собакой слабого типа и собакой сильной вариации слабого типа… От случайного скрещивания (V) Дедди с неизвестным самцом родился потомок под кличкой Орт. Несмотря на отсутствие пассивно-оборонительной реакции, Орт оказался слабого типа. При скрещивании (III) со своей сестрой Мелиной, родившейся от той же матери, но другого отца (см. скрещивание I этого семейства), были получены шесть потомков. Все они оказались с пассивно-оборонительной реакцией (один из них Валка — слабый тип сильной вариации). При скрещивании (IV) Орта со своей дочерью Мирри родилось пять потомков с пассивно-оборонительной реакцией (типы нервной системы не определены). 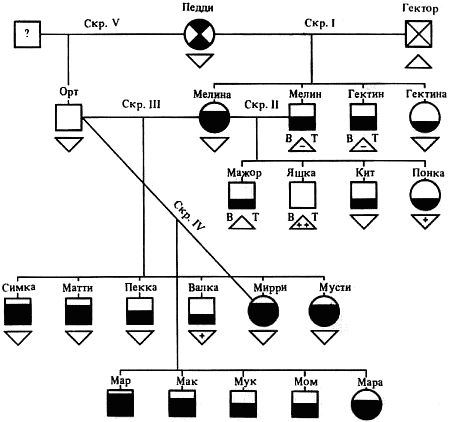 Рис. 4. Семейство № 4 Рис. 4. Семейство № 4 Семейство № 5 (рис. 5). В этом семействе был скрещен[17] (скрещивание I) нетрусливый холерик Курносый, у которого отставала подвижность, с сангвиником Найдой, у которой наблюдалось некоторое отставание тормозного процесса; ни у той, ни у другой собаки не было ни малейших следов пассивно-оборонительной реакции. Оба потомка, родившиеся от Найды, оказались сильными; один из них Наль — сильный сангвиник, другой Неля — сангвиник средней силы с отстающим тормозным процессом. Несмотря на сильный тип нервной системы, Наль проявил ясную пассивно-оборонительную реакцию (Тр-3). От скрещивания (III) между собой Нели и Наля был получен один потомок с пассивно-оборонительной реакцией (Тр-3) Нель-Наль — сангвиник слабой вариации. В этом семействе ясно выступает возникновение пассивно-оборонительной реакции независимо от типа нервной системы. 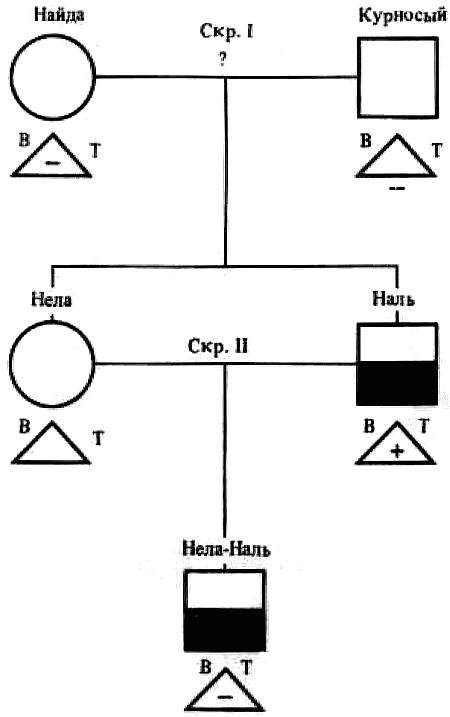 Рис. 5. Семейство № 5 Рис. 5. Семейство № 5 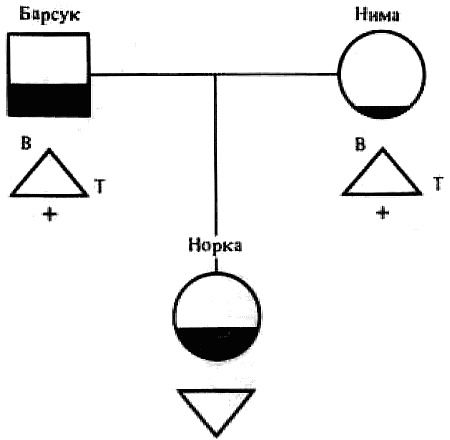 Рис. 6. Семейство № 6 Рис. 6. Семейство № 6 Семейство № 6 (рис. 6). В этом семействе были скрещены два холерика — Барсук и Нимфа. У обеих собак была хорошая подвижность раздражительного и тормозного процессов. Барсук проявил некоторую пассивно-оборонительную реакцию (Тр-2); у Нимфы были только незначительные следы оборонительного поведения. Единственный потомок, полученный от этого скрещивания (Норка), оказался слабого типа с наличием той же степени выражения трусливости (Тр-2), как у отца. Обобщая имеющиеся данные, можно сказать следующее. Фактический материал согласуется с приведенными выше данными о существенной роли генетических факторов в возникновении пассивно-оборонительной реакции у собак. Разделив условно всех обследованных нами собак на две группы: на собак с пассивно-оборонительной реакцией и без нее (считая собак с наличием пассивно-оборонительной реакции, начиная от Тр-2; собак же с наличием Тр-1 причисляя к группе нетрусливых собак), получаем сводную табл. 2. Таблица 2. Данные по наследование пассивно-оборонительной реакции (трусости) у собак * Помет Дедди и Гектора в этот материал не включен. Таблица 3. Данные по наследованию сильного и слабого типа собак Из этой таблицы ясно видна роль наследственности в возникновении пассивно-оборонительной реакции. Наибольший процент трусливых потомков оказывается в том случае, если у обоих родителей есть пассивно-оборонительная реакция, и наоборот, наименьший процент трусливых собак родится от нетрусливых родителей. Что наследственность играет значительную роль в возникновении пассивно-оборонительной реакции, видно также из корреляции, существующей между братьями и сестрами в отдельных пометах нашего материала: r = +0,54±0,06.[18] Так как все потомки, между которыми была вычислена корреляция, воспитывались в одних и тех же условиях, отнести ее за счет разных условий содержания едва ли возможно; сходство между братьями и сестрами в отдельных пометах должно было отнесено в основном за счет наследственных факторов. Таблица 4. Степень выражения пассивно-оборонительной реакции (трусости) у собак различных полов Перейдем теперь к вопросу о наследовании типологических свойств. Не вдаваясь в подробности обсуждения вопроса о характере и деталях наследования типологических свойств, укажем только, что приведенный материал с совершенной ясностью указывает на то, что типы нервной системы собак (во всяком случае сильный и слабый тип), как это и предполагал И. П. Павлов, наследуются. Это совершенно ясно видно как из приведенных родословных, так и из табл. 3, в которой весь материал разбит на две группы: собак сильного и слабого типа. Генетическая обусловленность типов видна также из корреляции, существующей между братьями и сестрами в отдельных пометах собак: r=+0,34±0,1.[19] На основании приведенных данных может даже создаться впечатление, что слабый тип наследуется в основном как рецессивный признак, однако недостаточность материала не позволяет сделать подобного вывода. Кроме того, мы полагаем, что монофакториальная схема едва ли может быть приложима к наследованию такого комплексного признака, как тип нервной системы; однако только дальнейшее накопление материала может окончательно уточнить этот вопрос. Прежде чем переходить к вопросу о зависимости между типом нервной системы и пассивно-оборонительной реакцией, остановимся на половом различии собак исследованного материала. В табл. 4 приведены данные по выражению пассивно-оборонительной реакции у собак различных полов. Как видно из этой таблицы и вычисленных средних (для самок М=2,39+0,32, для самцов М=2,17±0,29 — степени выражения трусости), статистически достоверной разницы в степени выражения пассивно-оборонительной реакции между самцами и самками не обнаруживается.[20] Таблица 5. Характеристика нервных процессов у собак различного пола В табл. 5 приведены данные о силе нервной системы собак различного пола. Для оценки связи между полом и типами нервной системы вычислен критерий соответствия (критерий Пирсона ?2). Он равен в данном случае X = 10,29. Вероятность соответствия P(?2), оказавшаяся меньше, чем 0,05, показывает, что разница между самцами по силе их нервной системы является достаточно существенной. Самцы обладают в среднем более сильной нервной системой, чем самки. Это вполне соответствует, как нам кажется, тем данным, которые получены Петровой при кастрации собак. Кастрация самцов, согласно ее опытам, приводит к ослаблению нервной системы собак. Возможно, несколько большая слабость самок по сравнению с самцами обусловлена отсутствием у них мужского полового гормона, который, очевидно, усиливает нервную систему собаки. Переходим теперь к вопросу о зависимости между типом нервной системы и оборонительным поведением собак. В большинстве семейств мы имеем ясную независимость возникновения и наследования типов нервной системы от трусости. Так, в семействе № 1 (скрещивание III) все собаки, у которых был определен тип, оказались сильными, но в то же время две из них (так же как и мать, которая была сильного типа, но трусливой) проявили ясную пассивно-оборонительную реакцию. В семействе № 2 весь помет проявляет в той или другой степени пассивно-оборонительную реакцию, однако в нем оказываются собаки как сильного, так и слабого типа. В семействе № 3 на фоне ясно выраженной трусости, проявляющейся у всех собак этого семейства, родятся собаки как сильного, так и слабого типа. Сходное явление наблюдается и в семействе № 4 (скрещивание III), в котором все трусливые потомки этого скрещивания оказываются как слабого, так и сильного типа. Таблица 6. Взаимоотношение между пассивно-оборонительной реакцией и типом нервной системы собак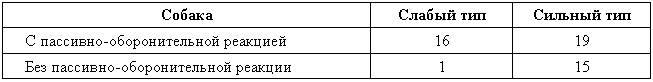 В семействе № 5 мы имеем, наоборот, рождение как трусливых, так и нетрусливых потомков, несмотря на то что все представители этого семейства сильного типа. Параллельное возникновение пассивно-оборонительного поведения и слабого типа мы имеем в семействе № 4 (скрещивание IV), в котором все потомки оказались слабыми и трусливыми, несмотря на то что их отец (слабого типа) не обладал пассивно-оборонительной реакцией. Некоторый намек на параллельное наследование слабого типа и пассивно-оборонительной реакции мы имеем и в скрещивании II этого семейства, в котором нетрусливый потомок, происшедший от этого скрещивания, оказался одновременно и наиболее сильным из всего помета. Коэффициент корреляции между пассивно-оборонительной реакцией и слабым типом собак изученной популяции r=+0,34±0,22 указывает на то, что статистическая связь между этими признаками не достоверна. Приближенная ошибка коэффициента корреляции в популяции, представители которой объединены в несколько родственных семей, должна вычисляться иначе, чем для свободно скрещивающейся. Вычисление ошибки произведено по формуле, предложенной М. В. Игнатьевым, за что считаем приятным долгом выразить ему свою благодарность. При пользовании этим методом, ошибка коэффициента корреляции вычисляется по формуле: 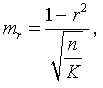 при чем величина K вычисляется по формуле: 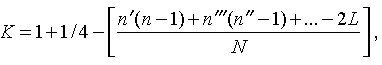 где n' — число особей в первом семействе; n' — число особей во втором семействе; n' — число особей в третьем семействе и т. д.; L — число семей; N — сумма всех обрабатываемых особей; 1/4 — минимальная степень корреляции, ожидаемая внутри отдельных семейств в обрабатываемом нами материале и возникающая в результате общего происхождения потомков внутри отдельных семейств. Необходимость такого вычисления диктуется тем, что в популяции, в которой имеется несколько семей, ошибка должна быть увеличена, так как наследственное сходство между родственниками уменьшает число независимых (случайных) наблюдений. Величина К и является тем коэффициентом, который вносит эту поправку. В табл. 6 приведены данные, показывающие частоту сочетания пассивно-оборонительной реакции с сильным и слабым типом. Как видно из этой таблицы, собаки с пассивно-оборонительной реакцией могут быть столь же часто как слабого, так и сильного типа. Собаки без пассивно-оборонительной реакции в подавляющем большинстве случаев оказались сильного типа. Наличие пассивно-оборонительной реакции у собак всех изученных типов ясно свидетельствует о том, что связи между пассивно-оборонительной реакцией и слабым типом нервной системы нет. Однако тот факт, что подавляющее большинство собак без пассивно-оборонительной реакции оказалось сильного типа, требует анализа этого явления. Возможно дать этому два объяснения. Во-первых, можно предположить, что пассивно-оборонительная реакция, не являясь непосредственным свойством слабого типа (против этого говорит отсутствие трусости у Орта, собаки слабого типа), все же более легко возникает у слабых собак. Во-вторых, можно предположить, что слабый тип чисто случайно сочетается большее количество раз с пассивно-оборонительной реакцией, чем с ее отсутствием. За это говорят следующие данные: если внимательно присмотреться к приведенным родословным, то в большинстве из них в силу случайного подбора скрещиваний трудно и ожидать сочетания слабого типа с отсутствием пассивно-оборонительной реакции. Так, в семействе № 1, в котором родится наибольший процент нетрусливых собак, родители оказались сильными, поэтому большинство потомков должно быть сильного типа. То же относится к семейству № 5. В семействах № 2 и № 3 хотя и встречаются слабые собаки, однако все как сильного, так и слабого типа потомки возникают на фоне пассивно-оборонительной реакции, распространенной в этих семействах. Едва ли есть какие-нибудь основания ожидать, чтобы слабые представители этих семейств были бы нетрусливыми. То же относится и к скрещиванию I семейства № 4. Мы считаем, что рождения слабых и нетрусливых собак можно было ожидать наиболее вероятно в скрещивании II и III семейства № 4. В скрещивании II этого семейства произошло расщепление по типу нервной системы и оборонительной реакции так, что в этом помете можно было ожидать рождения нетрусливых, но слабых собак. В скрещивании III этого семейства все потомки от двух слабых родителей оказались слабыми. При независимом наследовании и возникновении слабого типа и пассивно-оборонительной реакции надо было ожидать рождения некоторого числа нетрусливых потомков.[21] Этого не произошло. Таким образом, на основании того, что среди потомков в этих двух пометах не встречается собак с комбинацией слабый тип — отсутствие трусости, возникают некоторые основания ожидать существования отрицательной зависимости между слабым типом и отсутствием пассивно-оборонительной реакции. Однако на основании двух этих пометов утверждать о наличии такой зависимости едва ли возможно. Таким образом, подводя итог всему изложенному выше, можно сказать следующее: приведенный материал с несомненностью подтверждает данные предыдущих работ (Крушинский) о генетической обусловленности пассивно-оборонительной реакции у собак. Подтверждается мнение Павлова об очевидной наследственной обусловленности типов нервной системы собак (во всяком случае сильного и слабого типов). Анализ параллельного возникновения пассивно-оборонительной реакции и типов нервной системы указывает, что пассивно оборонительная реакция может свободно комбинироваться как с сильным, так и со слабым типом нервной системы. Что касается собак без пассивно-оборонительной реакции, то это были в подавляющем большинстве собаки сильного типа. Является ли это общей закономерностью или результатом случайного подбора скрещиваний на основании имеющегося в нашем распоряжении материала, ответить нельзя. Анализ показал, что по степени выражения пассивно-оборонительной реакции статистически достоверной разницы между самцами и самками обнаружить не удается. В то же время разница по степени силы нервной системы оказалась достаточно достоверной. Все это приводит нас к выводу, что пассивно-оборонительная реакция и тип высшей нервной деятельности являются в значительной степени самостоятельными свойствами нервной системы, которые могут в значительной степени независимо комбинироваться. Так, условия воспитания сильно влияют на оборонительную реакцию, однако, видимо, сравнительно слабо отражаются на общей характеристике типа нервной системы. Это указывает на разные физиологические процессы, лежащие в основе данных свойств поведения собак. Заключение Проведенный анализ наследования пассивно-оборонительного поведения собак в связи с типом нервной системы показал значительную независимость возникновения оборонительного поведения от типа нервной системы. Этот вывод полностью подтверждает мнение, высказанное И. П. Павловым в последние годы его жизни, что трусость собак не отождествима со слабым типом. В этом направлении важные данные были получены, как мы уже указывали, Майоровым, воспитывавшим щенят в условиях изоляции и показавшим огромное значение воспитания для формирования оборонительного поведения у собак. Однако воспитание является только одной из причин окончательного формирования типологических и темпераментных особенностей поведения взрослого животного. Другая сторона, определяющая поведение животного, — это наследственность. Работа Майорова дала материал для анализа только одного из путей связи между оборонительными и типологическими особенностями поведения — было исследовано влияние воспитания. Другим путем, ведущим к той же цели — выяснению существующих взаимоотношений между отдельными компонентами поведения, — является наследственный анализ. Этот метод дает возможность проникнуть в сущность тех или других особенностей поведения животного, ибо позволяет четко отдифференцировать отдельные врожденные реакции поведения и рассмотреть их взаимодействие в различных соотношениях в свете истории их наследственного возникновения. В настоящей работе была произведена попытка проанализировать связь между пассивно-оборонительной реакцией и типом нервной системы, пользуясь генетическим методом. Этот метод дал возможность проследить историю наследственного происхождения оборонительных и типологических особенностей поведения собак, указав на очевидную независимость их возникновения. Взгляд И. П. Павлова, высказанный им на основании работы Майорова, полностью подтвердился при анализе наследования свойств поведения. Анализ наследования свойств поведения является, таким образом, новым методом, который поможет физиологу расчленять сложное поведение животных на отдельные компоненты и рассматривать их взаимодействие в различных соотношениях. Являясь методом, который поможет изучению закономерностей физиологии нервной системы, наследственный анализ свойств поведения интересен еще и потому, что может пролить свет на вопрос о связи между функциональными и морфологическими свойствами организма. Единство формы и функции указывает на необходимость ожидать, что в основе наследования всякого признака поведения должно лежать наследование каких-либо морфологических или химических свойств организма, уже на основе которых и развивается данное поведение. Установление такой связи поможет изучению этапов онтогенетического возникновения признаков поведения, поможет изучению динамики их развития. Примечания:1 Биол. журн. 1938. Т. 7, № 4. С. 869–892. 2 Возможно, наследуется не пассивно-оборонительная реакция собак, а слабый тип нервной системы, и лишь на его фоне развивается пассивно-оборонительное поведение. 8 Слово «трусость» нами используется в качестве технического термина. Трусостью мы обозначаем пассивно-оборонительную реакцию. 9 Тр. Ин-та эволюц. физиологии и патологии высш. нерв. деятельности им. И. П. Павлова. 1947. Т. 1. С. 39–62. 10 Возможно эта склонность обусловлена свойством быстрой выработки условных рефлексов, так как можно думать, что секундирование является условным рефлексом, причем условным раздражителем является вид собаки, стоящей на стойке, а раздражителем, вызывающим безусловный рефлекс, — запах птицы. Очевидно, сочетание этих раздражителей привело к тому, что собака начала делать стойку не только на запах птицы, но и при виде собаки, стоящей на стойке. Можно предположить также, что эта склонность связана с имитацией, так как в последние годы появились данные, ясно указывающие на роль подражания в поведении животных. (Штодин, 1941). 11 Тр — трусость; цифровой показатель характеризует степень выражения этого признака. 12 Иногда встречаются собаки, которые проявляют в первый момент приближения к ним незначительные следы испуга (опускают хвост, отходят на 1–2 шага от решетки). Однако при заходе в клетку и при звуковом раздражителе не проявляют испуга. Таких собак мы обозначаем Тр-1/2, считая их разновидностью собак Тр-0. 13 Стандарт испытания напечатан в тезисах доклада Т. А. Тимофеевой и др., сделанного на IX Совещании по физиологическим проблемам в марте 1941 г. 14 Пробанаконцентрацию тормозного процесса. 15 Оценка оборонительного поведения для каждой собаки по результатам его определения. 16 Оборонительные реакции Дедди и Гектора не были определены, так как собаки давно пали; сведения об их оборонительном поведении получены от сотрудников Института, хорошо помнивших этих собак; кроме того, оборонительное поведение Дедди описано в работе Строгой. При дальнейшей обработке материала по оборонительным реакциям эти собаки в обрабатываемый материал не включались. 17 Это скрещивание не вполне достоверно, так как Найда была куплена уже беременной и хозяин утверждал, что она скрещена с Курносым, который был куплен в том же месте. 18 Так как популяция, в которой вычислялась корреляция, имеет большое количество родственников, ошибка должна быть несколько больше, чем в свободно скрещивающейся популяции. Это относится ко всем случаям вычисления ошибки в данной популяции. Однако это обстоятельство имеет практическое значение для данных, приведенных ниже. 19 Несколько меньший коэффициент корреляции по сравнению с таковым для пассивно-оборонительной реакции еще не означает существенного отличия в степени генетической обусловленности типологических свойств; это может быть вызвано меньшей градуированностью самого признака или несколько иным способом его наследования (более сильной доминантностью). 20 После того как была написана эта статья, автором был собран большой материал, главным образом на служебных собаках, показавший, что пассивно-оборонительная реакция резче выражена у самок (Крушинский, 1946). 21 Рождения нетрусливых собак от этого скрещивания можно ожидать по той причине, что Мелина при скрещивании даже с трусливым самцом дала одного нетрусливого потомка; тем более было основание ожидать рождения нетрусливых особей при скрещивании с нетрусливым Ортом. |
|
||